

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Summary
Neuropilin is a type I transmembrane protein and the molecular mass is 120 kDa. Two homologues, Neuropilin-1 and -2, are identified. The primary structure of Neuropilin-1 and Neuropilin-2 is well conserved and is divided into four domains, CUB (a1/a2) domain, FV/FVIII (b1/b2) domain, MAM (c) domain, and (d) domain that contains a transmembrane and a short cytoplasmic region. Both Neuropilin-1 and Neuropilin-2 have truncated and secreted form of splice variants. Neuropilins act as a receptor for two different extracellular ligands, class 3 semaphorins and specific isoforms of vascular endothelial growth factor. In both cases, neuropilin requires an additional transmembrane molecule to exhibit biological activity. Plexin-A is essential for class 3 semaphorin signaling. Vascular endothelial cell growth factor (VEGF) receptor is the major receptor for VEGF and neuropilin acts as isoform specific co-receptor for VEGF. The CUB and FV/FVIII domains of Neuropilin are the binding sites of semaphorin and VEGF. The MAM domain mediates semaphorin signaling to Plexin-A. Cross talk between semaphorin and VEGF on neuropilin suggests that class 3 semaphorins and the secreted forms of neuropilin act as antagonists to VEGF and its related growth factors.
Introduction
Neuropilin (NRP) is a single-spanning membrane protein and the molecular mass is 120 kDa. The protein has been firstly identified as an antigen of a specific antibody A5, which recognized the developmental stage of Xenopus optic nerve.1 The protein is unique to vertebrates, and so far, zebra fish, frog, chick, mouse, rat and human NRP homologues have been partially or completely identified. No homologous proteins in invertebrates have been reported. The expression pattern of NRP varies among species. Based on the expression pattern and stage in chick2 and mice,3 it has been speculated that NRP is involved in the formation of nervous system. However, the role of NRP had not been exactly revealed until two ligands were identified.
In 1997, two groups independently reported that NRP is the receptor for Sema3A, one of the class 3 semaphorin.4,5 Since an orthologue of NRP was reported as Neuropilin-2 (NRP2) at that time, NRP was renamed as Neuropilin-1 (NRP1). Sema3A is one of the members of Semaphorin family that is involved in the axon guidance during embryonic developmental stages.6 The application of anti-NRP1 antibody blocked Sema3A-induced growth cone collapse of rat E14 dorsal root ganglion (DRG) cells, confirming that the protein acted as a functional receptor for Sema3A. This fact is further strengthened by mutant mouse studies. DRG growth cones of NRP1−/− mice did not respond to Sema3A.7 Both Sema3A−/− and NRP1−/− mutant mice exhibited similar phenotype in nervous systems, such as aberrant and defasciculated peripheral nerve projection.7,8
In 1998, a different aspect of NRP1 was revealed. NRP1 also acts as an isoform specific receptor for Vascular Endothelial cell Growth Factor (VEGF).9 VEGF is a growth factor that stimulates the migration and proliferation of endothelial cells.10,11 One of the major isoforms of VEGF, VEGF165, binds to NRP1. The biological role of NRP1 in vascular system is proved by the studies of NRP1−/− mutant mice and NRP1 transgenic mice.12,13 Both NRP1−/− and transgenic mice died before birth and the vascular regression in the NRP1−/− embryos was in marked contrast to the overproduction of vessels in the embryos transgenic NRP1.
A unique character of NRP is that the protein is unable to generate the intracellular signaling. Instead, additional molecules are required to exhibit the biological function of Semaphorin and VEGF. For semaphorin signaling, Plexin-A (Plex-A) acts as an essential signal transducer.14 For VEGF signaling, VEGFR1(flt-1), or VEGFR2 (KDR/flk-1) are the major and functional receptor molecule and NRP serves as modulator.9
This section describes the structure of NRP1 and NRP2 at genome and protein level, then discusses the relation of the structure and the biological function of NRPs.
Primary structure and genomic structure of Neuropilin
Both NRP1 and NRP2 have transmembrane and truncated forms.15 The transmembrane forms of NRP1 and NRP2 share similar primary structure (Fig. 1). Following a short stretch of secretion signal, NRP1 and NRP2 consist of four different domains, two repeats of CUB domain (a1/a2), two repeats of FV/VIII domain (b1/b2), a MAM (c) domain, and a fourth domain (d) that contains transmembrane and relatively short 40 to 43 amino acid cytoplasmic region. The first CUB domains have significant homology with complement factor C1s/C1r, Bone Morphogenetic Protein 1(BMP1), and Tolloid proteins. The second FV/VIII domain shares the homology with coagulation factor FV/VIII, one of the receptor type tyrosine kinase DDR, and discoidin-1. The third domain MAM is the abbreviation of meprin, A5 (former name of NRP), and receptor protein-tyrosine phosphatase mu and kappa (PTPm, k).
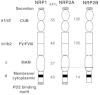
Full-length NRP2 has two major alternate splice variants, NRP2A and NRP2B.16 While NRP1 and NRP2A show 44% amino acid identity in entire regions, the fourth domain of NRP2B is unrelated to NRP1 or NRP2A. The amino acid identity of each domain, CUB, FV/FVIII, and MAM is 45%, 48% and 35%, respectively. In the fourth domain, NRP1 and NRP2A shares 49% identity, while NRP1 and NRP2B shares only 15% identity. Although the fourth domain has no apparent homology with other proteins, NRP1 and NRP2A possess a PSD-95/DIg/ZO-1 (PDZ) binding motif at the carboxyl termini. It has been shown that a Neuropilin-1 Interacting Protein (NIP) is associated with the NRP1 carboxyl terminus.17
The completion of human genome sequence18 and a report from Klagsbrun laboratory15 prompted us to examine the genomic structure of NRPs (Fig. 2). The loci of human NRP1 and NRP2 genes are 10p12 and 2q34, respectively. The loci of mouse versions are chromosomes 8 and 1. Human NRP1 gene spans a length of approximately 157 kb and is split into 19 exons. Human NRP2 is 115 kb length and is divided into 23 exons. The exon and intron numbers in this section are according to the assignment of the Human Genome Sequence.18 Some of the exons within the contig are not used for encoding NRPs. These exons are shown as white half size boxes in Fig 2. The exons 3 and 9 in NRP1 locus generate different mRNAs. NRP2 locus contains three exons (5, 6, 21) unrelated to encode NRP2 mRNA. The relation between these exotic exons and NRP genes is currently unknown.

The location of exon-intron junction is similar for these two genes. Fourteen of the 16 splice sites are conserved between NRP1 and NRP2 gene. These sites are found in the exons encoding CUB and FV/FVIII domains, correlating well with the amino acid homologies of these domains. The splicing points corresponding to the regions of MAM and transmembrane are less conserved. The similar structure of NRP1 and NRP2 suggests a duplication of these genes in the evolution of vertebrates.
Alternative splice variants of full-length NRP2, NRP2A, and NRP2B are generated by the splicing of the exons 19, 20, and 22. The fourth domain of NRP2A and NRP2B is encoded by exon 22 and exon 20, respectively. Comparing NRP2A and NRP2B, NRP2A is 17 amino acid length longer than NRP2B between the junction point and transmembrane region. Another alternate splicing in exon 19 generates 5 amino acid insertion flanked with the sequence encoded by exon 20 or exon 22. Then four splice variants, NRP2A17, NRP2A22, NRP2B0, and NRP2B5 are generated. In mice, two additional variants, NRP2A0 and NRP2A5, were also reported.16 NRP2A and NRP2B show slightly different expression patterns in adult humans.15 NRP2A is predominantly expressed in liver, placenta, lung, intestine, heart and kidney, while NRP2B is found in heart and skeletal muscle. Significant level of expression of both forms is detectable in adult brain. The role of structural difference between NRP2A and NRP2B is not clear. However, since the carboxyl termini of NRP2B variants are not homologous to the termini of NRP1 and NRP2A, NRP2B may not bind to NIP.
It has been identified two truncated forms of NRP1, s11NRP1 and s12NRP1, and one short form of NRP2, s9NRP2.15,19 All of the truncated forms are generated by the use of alternate polyadenylation signals in the specific introns. Within the mRNA of s11NRP1, the sequence of exon 13 is flanked with a 1866 base intron 13-derived sequence encoding a 84 unique amino acid sequence. The mRNA of s12NRP1 has a 28 base intron 14-derived sequence after the exon 14 junction. Since exon 13 and 14 encode FV/FVIII domain, s11NRP1 and s12NRP1 consist of only CUB and FV/FVIII domains. No apparent hydrophobic regions are encoded by the intron-derived sequences. Then, two truncated forms of NRP1 are secreted proteins. These variants are predominantly expressed in placenta, liver, heart, kidney and lung. The expression of both forms in brain is lower than other tissues.
A soluble form of NRP2, s9NRP2, is also generated by the same manner as the truncated forms of NRP1.15 The mRNA of s9NRP2 is flanked with 144 bp intron 13-derived sequence after exon 13. The 144-bp sequence contains a stop codon and a polyadenylation site. Then s9NRP2 consists of two CUB domains, the first b1 of FV/FVIII domain, a part of b2 domain, and a 8 unique amino acid sequence encoded by the intron 9.
Binding properties of NRP domains
Semaphorins are a large family of transmembrane and secreted proteins are mainly involved in axon guidance.6 All of the semaphorins possess a 550-amino acid Semaphorin (Sema) domain at their amino termini and are divided into 8 classes based on species, amino acid sequence, and structural similarity. Classes 1 and 2 are invertebrate semaphorins, classes 3 to 7 are vertebrates, and class V is viral semaphorins. Classes 1, 4, 5, 6 and 7 are membrane bound proteins, while classes 2, 3 and V are secreted semaphorins.
So far, at least three different types of binding proteins were reported as semaphorin receptors, Neuropilin,4,5 Plexin,20 and CD72.21 Classes 1 and 2 invertebrate semaphorins bind to Plexin.22 In vertebrates, class 4, 5, 6, 7 semaphorins bind to Plexin B, C or A directly, and class V semaphorins bind to Plexin C.23 Plexins possess a Sema domain at the amino termini, suggesting that the proteins are distant ancestors of semaphorins. It is interesting to note that CD72 serves as a functional receptor for Sema4D (CD100) in lymphocytes.21 CD72 is a 45 kDa type II membrane protein that belongs to the C-type lectin. Sema4D binds to CD72 with a Kd of 300 nM and augments the effect of CD40 on B cell responses, such as proliferation.
A unique feature of class 3 semaphorins is that they bind to NRPs but not to plexins. Six members of semaphorins, 3A, 3B, 3C, 3D, 3E and 3F, belong to class 3. This class of secreted protein consists of a Sema domain, one immunoglobulin domain and a basic amino acid rich region. Class 3 Semaphorins form a homodimer through a disulfide bond in the basic rich region, which is critical to exhibit biological activity.24
NRP1 and NRP2 exhibits different specificity to class 3 semaphorins. NRP1 binds to all class 3 semaphorins while NRP2 binds to Sema3B, Sema3C, Sema3D, Sema3E, and Sema3F but not to Sema3A.25 Although NRP1 and NRP2 display relatively large spectrum of binding to class 3 semaphorins, this does not account for the specific effect of each member of class 3 semaphorins. For example, the DRG express only NRP1 and the growth cones are sensitive to Sema3A but not Sema3B or 3C, while sympathetic ganglion cells that express both NRP1 and NRP2 are repelled by Sema3A, Sema3B, and Sema3C.26
The characterization of class 3 semaphorin binding to NRP1 and NRP2 has been conducted on Sema3A, 3C and 3F. He and Tessier-Lavigne showed that Sema3A binds to NRP1 with two different regions, the Sema domain and the basic rich region.4 The broad binding specificity of NRP1 reflects the binding of the basic rich region of class 3 semaphorins. For example, full-length Sema3F binds to NRP1 and NRP2, while Sema-Ig domain of Sema3F shows higher affinity to NRP2. In contrast, the basic rich region of Sema3F binds effectively NRP1 but not NRP2.27 The discrepancy of binding specificity between Sema-Ig domain and basic rich region compromises broad spectrum of class 3 semaphorin binding to NRPs.
However, the binding of Sema domain to NRPs, but not of basic rich region, is the initial step to elicit the biological function of class 3 semaphorins. The growth cone collapsing activity is retained in a chimera protein consisting of the Sema domain of Sema3A fused to the Fc region of human IgG1.28 The importance of Sema domain is also supported by the study of chimera of chick Sema3A (Collapsin-1) and Sema3D (Collapsin-2),29 in which Sema3A but not Sema3D induced growth cone collapse of embryonic chick DRG cells. In this study, swapping of 70 amino acid region of Sema3A and Sema3D within the Sema domain altered the growth cone collapse activity of both proteins. Each member of class 3 semaphorin uses different combinations of NRP1 and NRP2 to exhibit repulsive action. For instance, Sema3A action is mediated by NRP1 whereas Sema3F is mediated by NRP2. On the other hand, the action of Sema3C requires both NRP1 and NRP2.27
Five different groups performed deletion and chimera analyses on NRP1 and NRP2.27,30–33 Nakamura et al30 and Chen et al27 demonstrated that the CUB domains of NRPs are the binding site for Sema domain of class 3 semaphorins. A deletion mutant NRP1Δ 276–797 that contains only the CUB domain of NRP1 was able to bind the Sema-Ig portion of Sema3A (Fig. 3). The CUB domain also determines the binding preference of NRP2. A NRP1/NRP2 chimera 2111 in which CUB domain of NRP1 was substituted with the one of NRP2 bound to Sema3C but not to Sema3A.30 The selective binding of 2111 chimera to the semaphorins was similar to the specificity of NRP2. These results indicate that the CUB domain is the primary binding site of the Sema domain. In contrast, the basic rich region of Sema3A was bound to the boundary of CUB and FV/FVIII domains. The basic rich region bound to NRP1Δ 18–253 (Fig. 3)31 but not to NRP1Δ 18–282.30 This suggests that the region including a 27 amino acid stretch from 255 to 282 of NRP1 is critical for the binding of the basic rich region. It has been shown that the FV/FVIII domain of NRP1 is involved in the binding of Vascular Endothelial cell Growth Factor (VEGF)31 and in NRP1-mediated cell adhesion.33

While the CUB and FV/FVIII domains are involved in the binding of class 3 semaphorin and other ligands, the MAM domain participates in the signal transmission of semaphorins. The MAM domain of NRP shares homology with receptor protein tyrosine phosphatase m and meprin. It has been shown that the MAM domain participates in the oligomerization of NRPs.30
The functional role of the MAM domain in Class 3 semaphorin signaling was demonstrated by Sema3A responsiveness of the chick retinal ganglion cells engineered to express full-length NRP1 or a series of NRP1 deletion mutant. Chick retinal ganglion cells lack normally NRP1 expression, therefore the growth cones do not respond to Sema3A. Herpes Simplex Virus vector-mediated expression of NRP1 in these cells renders Sema3A responsiveness. Using this system, series of deletion and chimera mutants of NRP1 were introduced and examined.30 The experiments showed that the MAM domain deleted mutant was unable to transmit Sema3A signaling. This was consistent with other studies: a MAM domain deleted NRP1 mutant also showed a dominant-negative effect on Sema3A-induced growth cone collapse of embryonic chick sympathetic neurons.32 The antibody directed against the MAM domain of NRP2 blocked the Sema3F-induced growth cone collapse of sympathetic ganglion cells.31 These results indicates that the MAM domain is the interface of semaphorin signaling. Then, what kind of molecule interacts with the MAM domain and acts as a signal transducer?
Neuropilin-1 Interacting Protein binds to the carboxyl terminus of NRP1
Shortly after the identification of NRP1 as a receptor for Sema3A, efforts to identify the downstream of Sema3A signaling began. A straightforward approach was the use of Yeast two hybrid system for searching binding partners of the intracellular region of NRP1. This method successfully isolated Neuropilin-1 Interacting Protein (NIP), which recognizes the four amino acids of carboxyl terminus of NRP1 (Figs. 1 and 4).17 NIP is a 40-kDa protein and possesses one central domain that shares significant homology with PSD-95/Dig/ZO-1(PDZ). It has been shown that this domain binds to the four amino acid sequence of NRP1 carboxyl terminus, Tyr-Ser-Glu-Ala. While NRP2A also contains a similar sequence Cys-Ser-Glu-Ala at the carboxyl terminus, the carboxyl terminus of NRP2B is different from NRP1 and NRP2A. The deletion of three amino acids from the carboxyl terminus of NRP1 diminishes the interaction with NIP.
NIP has been independently cloned as RGS-GAIP-interacting protein (GIPC)34 and SEMCAP-1.35 RGS-GAIP is a GTPase-activating protein (GAP) for Gαi subunits, which is localized on clathrin-coated vesicles. The carboxyl terminus of RGS-GAIP is Ser-Glu-Ala, which exactly matches to NRP1. SEMCAP-1 was identified as an interacting protein with the intracellular region of Sema4C (M-SemaF). The carboxyl terminus of Sema4C is Glu-Ser-Ser-Val, limiting homology to the termini of NRP1 and RGS-GAIP. Then, X-Ser-X-Ala/Val is the consensus motif of NIP/GAIP/SEMCAP-1 binding site. Since NIP is a cytoplasmic protein and the extracellular domain of NRP transmits the Semaphorin signal to another protein, the involvement of NIP in class 3 semaphorin signaling has not been well defined.
Additional receptor required for signal transduction
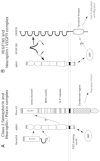
As described in Chapter 5, plexin is the receptor for certain class of semaphorins. Drosophila Sema1a interacts with Drosophila Plex-A.22 Class 7 semaphorins and viral Semaphorins bind to Plex-C.20,23 Class 3 semaphorins do not bind plexins directly, however, Plexin-A forms complex with NRP and acts as signaling molecule.14 This is supported by the following facts. While COS-7 cells expressing NRP1 bound Sema3A with a Kd of 1 nM, Plex-A1 co-expression increased the affinity of NRP1 to Sema3A about five-fold with a Kd of 0.2 nM. Furthermore, COS-7 cells co-expressing NRP1 and Plex-A1 presented contracted cell morphology after Sema3A exposure. This phenomenon resembles to the collapsed state of growth cones. Interestingly, the cells expressing MAM domain deleted NRP1 and Plex-A1 did not exhibit high affinity binding site for Sema3A nor alter the Figure 4. Schematic representation of Neuropilin and its interacting proteins morphology after Sema3A stimulation. This fact clearly demonstrates that Sema3A-collapsing signal is transmitted from MAM domain to Plex-A1. Finally, the overexpression of a mutant Plex-A1 without intra-cytoplasmic domain blocked Sema3A-induced growth cone collapse of chick DRG. This indicates that the Plex-A1 mutant acts as dominant negative manner and the cytoplasmic tail of Plex-A1 initiates an intracellular signal of collapse response. Other members of Plexin-A, Plex-A2 and Plex-A3 have been shown to mediate class 3 semaphorin signals.36,37
Vascular endothelial cell growth factor (VEGF-A) is a potent factor that induces the formation of blood vessels.10 VEGF forms a 40-45 K homodimer and has low homology with platelet-derived growth factor. Five different polypeptides, 121, 145, 165, 189 and 206 amino acids (VEGF121-VEGF206) are generated by alternative splicing from VEGF-A gene, each of them capable of making an active homodimer. VEGF121 and VEGF165 are the most abundant forms. A unique 44 amino acid sequence of VEGF165 is derived from exon 7. VEGF121 possesses the biological activity of VEGF, the stimulation of proliferation and migration of endothelial cells. Comparing to VEGF121, VEGF165 is a more potent mitogen for endothelial cells, suggesting the modulating role of the unique sequence of VEGF165.
At least two receptor type tyrosine kinases, VEGFR1 (Flt-1) and VEGFR2 (KDR/Flk-1), serve as VEGF receptors (Fig. 4).10 Both VEGFR1 and VEGFR2 have 7 immunoglobulin repeats in the extracellular region, a transmembrane domain, and an intracellular tyrosine kinase domain. The biological activity of VEGF is exhibited through the dimerization of the receptor, subsequently leading to the activation of the tyrosine kinase.
Besides these main receptors, Soker et al9 reported that NRP1 is a specific receptor for VEGF165 but not for VEGF121. This finding is quite consistent with the expression pattern of NRP1 as well as mutant NRP1 mice phenotype.12 The interactions between VEGFR and NRP are fully detailed in Chapter 7.
Porcine aortic endothelial (PAE) cells lack expression of NRP1 and VEGFR2, allowing to the expression and functional study of these receptors in vascular cells. When NRP1 was expressed in PAE cells, VEGF165 bound to NRP1 with a Kd of 0.3 nM. While VEGF stimulated the migration of PAE cells expressing VEGFR2, VEGF165 could not alter the migration of PAE cells expressing NRP1. However, co-expression of NRP1 and VEGFR2 in PAE cells augmented the migration upon the stimulation of VEGF165, comparing to single-expression of VEGFR2. The direct interaction of NRP1 and VEGFR2 was also shown by co-immunoprecipitation.38 These results demonstrate that NRP1 acts as a co-receptor of VEGF165. NRP2 also serves as co-receptor for VEGF145 as well as for VEGF165. Other related homologues of VEGF, placenta growth factor-2 (PlGF-2) have been shown to bind NRP1 and NRP2.
Although the function of NIP in Class 3 semaphorin signaling has not been demonstrated, NIP may participate in the signal transduction of VEGF. One hint has been provided by the study of the signal transduction of VEGF165 through NRP2 and VEGFR1 in PAE cells.39 While wild-type NRP2A could bind VEGF165, a tagged NRP2A containing a myc epitope at the carboxyl terminus, which disrupted the PDZ binding motif, could not bind to VEGF165. In contrast, Sema3F binding to NRP2A was not affected by the addition of myc tag. Considering the fact that NIP is a broadly expressed protein17,35 and it probably binds to the Ser-Glu-Ala motif of NRP2A carboxyl terminus, the homo- or hetero-oligomerization of NRP2A through NIP may be required for the binding of VEGF165.
It is of interest whether two distinct ligands may interfere with each other on one receptor molecule. The study using transient expression of NRP1 and VEGFR2 in COS-7 cells demonstrated that Sema3A inhibits the binding of VEGF165 to NRP1. Sema3A also antagonizes the VEGF165-induced migration of PAE cells co-expressing NRP1 and VEGFR2. Indeed, Semaphorin-NRP interaction plays positive or negative regulatory role in lung branching morphogenesis,40 and Sema3B and Sema3F have been implicated as tumor suppressor genes in human lung small cell carcinoma.41 These semaphorins may suppress the expansion of tumors by antagonizing VEGF and its related growth factors.
Bagnard et al42 reported that migration and apoptosis of neural progenitor cells was regulated by the balance of Sema3A and VEGF165. Furthermore, they observed that Sema3A activates the tyrosine kinase of VEGFR1. This suggests that VEGFR1 may serve as an additional component of semaphorin receptor, at least during the migration stage of neural progenitor. Further investigation is required to prove this idea.
As mentioned earlier, soluble forms of NRP1 and NRP2 are predominantly expressed in non-neuronal tissues. These truncated forms of NRP may also act as an inhibitor of VEGF-induced vascular formation. When rat prostate carcinoma-derived cell lines were injected to a rat host, tumor masses were formed in various organs. These masses were invaded by numerous blood vessels because of the production of VEGF. When the same cell lines expressing soluble s12NRP1 were injected, most of the malignant cells in the mass were destined to apoptotic degradation.19 Poor formation of blood vessels in the tumors was also observed. In this case, the s12NRP1 probably antagonized VEGF action to malignant cells and to invading vascular endothelial cells. This opens the possibility of the therapeutic use of soluble NRPs as anti-tumor reagents.
Concluding remarks
Three distinct extracellular domains of NRP play important roles in semaphorin and VEGF signaling (Fig. 4). The CUB and FV/FVIII domains serve as the binding sites for the two ligands. The MAM domain acts as a signaling interface to Plex-A, at least in the case of class 3 semaphorin signal transduction (Fig. 4A). VEGF165 binds to both NRP and VEGFR (Fig. 4B). A PDZ protein, NIP binds to the carboxyl termini of NRP1 and NRP2A. NIP may play important role for VEGF signaling. Recent accumulating findings begin to resolve the mysterious action of class 3 semaphorins as tumor suppressor genes. Class 3 semaphorins seem to antagonize the binding of VEGF or VEGF related growth factors to NRPs. Then the Semaphorins regulate precisely the strength of these factors and maintain appropriate growth of various organs. The soluble forms of NRPs may also play a similar role by quenching the growth factors.
Now the role of NRPs as a multiple ligand receptor emerges. Furthermore, NRPs require an appropriate transmembrane molecule in accordance with the bound ligand to exhibit the biological function. As the signal transducer, class 3 semaphorins use Plex-A. L1, another cell surface molecule, is also thought to interact with NRP1 and to be involved in Sema3A signaling.43 In the case of VEGF, NRP acts as co-receptor of VEGFR. All of the molecules described in this Chapter, NRPs, Plex-A, L1, and VEGFR are single membrane spanning proteins and structurally unrelated or distant. This suggests that some of transmembrane proteins may form functional hetero-oligomers with other unrelated proteins rather than homo-oligomerization as seen in the activation of receptor tyrosine kinases. Verifying the known single membrane spanning proteins from this aspect may find alternate new role of those proteins.
Acknowledgments
We thank to Professor Stephen M. Strittmatter at Yale University for providing AP-Sema3A, Sema-Ig-AP, AP-basic, NRP1 and NRP1Δ 276–797(1001) expression vectors.
References
- 1.
- Takagi S, Hirata T, Agata K. et al. The A5 antigen, a candidate for the neuronal recognition molecule, has homologies to complement components and coagulation factors. Neuron. 1991;7(2):295–307. [PubMed: 1908252]
- 2.
- Takagi S, Kasuya Y, Shimizu M. et al. Expression of a cell adhesion molecule, neuropilin, in the developing chick nervous system. Dev Biol. 1995;170(1):207–222. [PubMed: 7601310]
- 3.
- Kawakami A, Kitsukawa T, Takagi S. et al. Developmentally regulated expression of a cell surface protein, neuropilin, in the mouse nervous system. J Neurobiol. 1996;29(1):1–17. [PubMed: 8748368]
- 4.
- He Z, Tessier-Lavigne M. Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell. 1997;90(4):739–751. [PubMed: 9288753]
- 5.
- Kolodkin AL, Levengood DV, Rowe EG. et al. Neuropilin is a semaphorin III receptor. Cell. 1997;90(4):753–762. [PubMed: 9288754]
- 6.
- Semaphorin Nomenclature Committee. Unified nomenclature for the semaphorins/collapsins. Semaphorin Nomenclature Committee. Cell. 1999;97(5):551–552. [PubMed: 10367884]
- 7.
- Kitsukawa T, Shimizu M, Sanbo M. et al. Neuropilin-semaphorin III/D-mediated chemorepulsive signals play a crucial role in peripheral nerve projection in mice. Neuron. 1997;19(5):995–1005. [PubMed: 9390514]
- 8.
- Taniguchi M, Yuasa S, Fujisawa H. et al. Disruption of semaphorin III/D gene causes severe abnormality in peripheral nerve projection. Neuron. 1997;19(3):519–530. [PubMed: 9331345]
- 9.
- Soker S, Takashima S, Miao HQ. et al. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell. 1998;92(6):735–745. [PubMed: 9529250]
- 10.
- Klagsbrun M, D'Amore PA. Vascular endothelial growth factor and its receptors. Cytokine Growth Factor Rev. 1996;7(3):259–270. [PubMed: 8971481]
- 11.
- Risau W. Mechanisms of angiogenesis. Nature. 1997;386(6626):671–674. [PubMed: 9109485]
- 12.
- Kawasaki T, Kitsukawa T, Bekku Y. et al. A requirement for neuropilin-1 in embryonic vessel formation. Development. 1999;126(21):4895–4902. [PubMed: 10518505]
- 13.
- Kitsukawa T, Shimono A, Kawakami A. et al. Overexpression of a membrane protein, neuropilin, in chimeric mice causes anomalies in the cardiovascular system, nervous system and limbs. Development. 1995;121(12):4309–4318. [PubMed: 8575331]
- 14.
- Takahashi T, Fournier A, Nakamura F. et al. Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors. Cell. 1999;99(1):59–69. [PubMed: 10520994]
- 15.
- Rossignol M, Gagnon ML, Klagsbrun M. Genomic organization of human neuropilin-1 and neuropilin-2 genes: identification and distribution of splice variants and soluble isoforms. Genomics. 2000;70(2):211–222. [PubMed: 11112349]
- 16.
- Chen H, Chedotal A, He Z. et al. Neuropilin-2, a novel member of the neuropilin family, is a high affinity receptor for the semaphorins Sema E and Sema IV but not Sema III. Neuron. 1997;19(3):547–559. [PubMed: 9331348]
- 17.
- Cai H, Reed RR. Cloning and characterization of neuropilin-1-interacting protein: a PSD-95/Dlg/ZO-1 domain-containing protein that interacts with the cytoplasmic domain of neuropilin-1. J Neurosci. 1999;19(15):6519–6527. [PMC free article: PMC6782790] [PubMed: 10414980]
- 18.
- International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature. 2001;409(6822):860–921. [PubMed: 11237011]
- 19.
- Gagnon ML, Bielenberg DR, Gechtman Z. et al. Identification of a natural soluble neuropilin-1 that binds vascular endothelial growth factor: In vivo expression and antitumor activity. Proc Natl Acad Sci USA. 2000;97(6):2573–2578. [PMC free article: PMC15970] [PubMed: 10688880]
- 20.
- Tamagnone L, Artigiani S, Chen H. et al. Plexins are a large family of receptors for transmembrane, secreted, and GPI-anchored semaphorins in vertebrates. Cell. 1999;99(1):71–80. [PubMed: 10520995]
- 21.
- Kumanogoh A, Watanabe C, Lee I. et al. Identification of CD72 as a lymphocyte receptor for the class IV semaphorin CD100: A novel mechanism for regulating B cell signaling. Immunity. 2000;13(5):621–631. [PubMed: 11114375]
- 22.
- Winberg ML, Noordermeer JN, Tamagnone L. et al. Plexin A is a neuronal semaphorin receptor that controls axon guidance. Cell. 1998;95(7):903–916. [PubMed: 9875845]
- 23.
- Comeau MR, Johnson R, DuBose RF. et al. A poxvirus-encoded semaphorin induces cytokine production from monocytes and binds to a novel cellular semaphorin receptor, VESPR. Immunity. 1998;8(4):473–482. [PubMed: 9586637]
- 24.
- Klostermann A, Lohrum M, Adams RH. et al. The chemorepulsive activity of the axonal guidance signal semaphorin D requires dimerization. J Biol Chem. 1998;273(13):7326–7331. [PubMed: 9516427]
- 25.
- Nakamura F, Kalb RG, Strittmatter SM. Molecular basis of semaphorin-mediated axon guidance. J Neurobiol. 2000;44(2):219–229. [PubMed: 10934324]
- 26.
- Takahashi T, Nakamura F, Jin Z. et al. Semaphorins A and E act as antagonists of neuropilin-1 and agonists of neuropilin-2 receptors. Nat Neurosci. 1998;1(6):487–493. [PubMed: 10196546]
- 27.
- Chen H, He Z, Bagri A. et al. Semaphorin-neuropilin interactions underlying sympathetic axon responses to class III semaphorins. Neuron. 1998;21(6):1283–1290. [PubMed: 9883722]
- 28.
- Eickholt BJ, Morrow R, Walsh FS. et al. Structural features of collapsin required for biological activity and distribution of binding sites in the developing chick. Mol Cell Neurosci. 1997;9(5-6):358–371. [PubMed: 9361274]
- 29.
- Feiner L, Koppel AM, Kobayashi H. et al. Secreted chick semaphorins bind recombinant neuropilin with similar affinities but bind different subsets of neurons in situ. Neuron. 1997;19(3):539–545. [PubMed: 9331347]
- 30.
- Nakamura F, Tanaka M, Takahashi T. et al. Neuropilin-1 extracellular domains mediate semaphorin D/III-induced growth cone collapse. Neuron. 1998;21(5):1093–1100. [PubMed: 9856464]
- 31.
- Giger RJ, Urquhart ER, Gillespie SK. et al. Neuropilin-2 is a receptor for semaphorin IV: Insight into the structural basis of receptor function and specificity. Neuron. 1998;21(5):1079–1092. [PubMed: 9856463]
- 32.
- Renzi MJ, Feiner L, Koppel AM. et al. A dominant negative receptor for specific secreted semaphorins is generated by deleting an extracellular domain from neuropilin-1. J Neurosci. 1999;19(18):7870–7880. [PMC free article: PMC6782461] [PubMed: 10479689]
- 33.
- Shimizu M, Murakami Y, Suto F. et al. Determination of cell adhesion sites of neuropilin-1. J Cell Biol. 2000;148(6):1283–1293. [PMC free article: PMC2174302] [PubMed: 10725340]
- 34.
- De Vries L, Lou X, Zhao G. et al. A PDZ domain containing protein, interacts specifically with the C terminus of RGS-GAIP. Proc Natl Acad Sci USA. 1998;95(21):12340–12345. [PMC free article: PMC22833] [PubMed: 9770488]
- 35.
- Wang LH, Kalb RG, Strittmatter SM. A PDZ protein regulates the distribution of the transmembrane semaphorin, M-SemF. J Biol Chem. 1999;274(20):14137–14146. [PubMed: 10318831]
- 36.
- Takahashi T, Strittmatter SM. PlexinA1 autoinhibition by the plexin sema domain. Neuron. 2001;29(2):429–439. [PubMed: 11239433]
- 37.
- Cheng HJ, Bagri A, Yaron A. et al. Plexin-a3 mediates semaphorin signaling and regulates the development of hippocampal axonal projections. Neuron. 2001;32(2):249–263. [PubMed: 11683995]
- 38.
- Whitaker GB, Limberg BJ, Rosenbaum JS. Vascular endothelial growth factor receptor-2 and neuropilin-1 form a receptor complex that is responsible for the differential signaling potency of VEGF(165) and VEGF(121). J Biol Chem. 2001;276(27):25520–25531. [PubMed: 11333271]
- 39.
- Gluzman-Poltorak Z, Cohen T, Shibuya M. et al. Vascular endothelial growth factor receptor-1 and neuropilin-2 form complexes. J Biol Chem. 2001;276(22):18688–18694. [PubMed: 11278319]
- 40.
- Kagoshima M, Ito T, Kitamura H. et al. Diverse gene expression and function of semaphorins in developing lung: positive and negative regulatory roles of semaphorins in lung branching morphogenesis. Genes Cells. 2001;6(6):559–571. [PubMed: 11442635]
- 41.
- Sekido Y, Bader S, Latif F. et al. Human semaphorins A(V) and IV reside in the 3p21.3 small cell lung cancer deletion region and demonstrate distinct expression patterns. Proc Natl Acad Sci USA. 1996;93(9):4120–4125. [PMC free article: PMC39497] [PubMed: 8633026]
- 42.
- Bagnard D, Vaillant C, Khuth ST. et al. Semaphorin 3A-vascular endothelial growth factor-165 balance mediates migration and apoptosis of neural progenitor cells by the recruitment of shared receptor. J Neurosci. 2001;21(10):3332–3341. [PMC free article: PMC6762465] [PubMed: 11331362]
- 43.
- Castellani V, Chedotal A, Schachner M. et al. Analysis of the L1-deficient mouse phenotype reveals cross-talk between Sema3A and L1 signaling pathways in axonal guidance. Neuron. 2000;27(2):237–249. [PubMed: 10985345]
- Structural and Functional Relation of Neuropilins - Madame Curie Bioscience Data...Structural and Functional Relation of Neuropilins - Madame Curie Bioscience Database
- Molecular Basis of Oncogenesis by NF-κB: From a Bird's Eye View to a RELevant Ro...Molecular Basis of Oncogenesis by NF-κB: From a Bird's Eye View to a RELevant Role in Cancer - Madame Curie Bioscience Database
- Preimplantation Genetic Diagnosis (PGD): Screening for Aneuploidy in Human Oocyt...Preimplantation Genetic Diagnosis (PGD): Screening for Aneuploidy in Human Oocytes and Polar Bodies - Madame Curie Bioscience Database
- Cytokines and Chemokines in Autoimmune Disease: An Overview - Madame Curie Biosc...Cytokines and Chemokines in Autoimmune Disease: An Overview - Madame Curie Bioscience Database
- Regulation of the Rod Photoreceptor Cyclic Nucleotide-Gated Channel - Madame Cur...Regulation of the Rod Photoreceptor Cyclic Nucleotide-Gated Channel - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...