

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Introduction
Serine protease inhibitors (serpins) are comprised of a large family of molecules that play a variety of physiological roles in vivo.13 Not all molecules that inhibit serine proteases are termed serpins. But all serpins have a very special protein structure and molecular weight about 400 amino acids.2 Serpins exist in almost every organism, from virus to mammals.35 They can be divided into two categories: inhibitory and non-inhibitory serpins. Non-inhibitory serpins, typified by ovalbumin and PEDF, do not exhibit protease inhibitor activity, but rather function as a storage protein and neural differentiation factor, respectively.6,7 Inhibitory serpins ablate serine proteases through their functional domain-reactive site loop (RSL).2 Interestingly, some inhibitory serpins have evolved other regulatory functions. For example, plasminogen activator inhibitor 1 (PAI-1) not only specifically inhibits tPA and uPA,8,9but also regulates cell adhesion, which is independent of its protease inhibitor function, by blocking integrin αvβ3 binding to vitronectin.10,11 This implies that serpins not only play diverse roles as a class, but also a single serpin molecule may possess multiple functions.12
Maspin is a unique member of the serpin family that shares extensive homology with monocyte-neutrophil elastase inhibitor,3 PAI-2 and other serpins.13,14 Initially identified as a class II tumor suppressor gene, maspin has been shown to inhibit invasion and motility of mammary tumors.13,15,16 Tumor transfectants expressing maspin exhibit decreased growth and metastasis in nude mice.13 Maspin gene expression is not detected in most breast tumors and loss of its expression is correlated with tumor invasiveness.17 Maspin is also found to be a potent angiogenesis inhibitor.18 In human breast tissue, maspin seems to be present more in luminal than myoepithelial cells, and it has been suggested that those maspin-expressing myoepithelial cells form a defensive barrier for the progression from ductal carcinoma in situ to more invasive carcinomas.19 A dispute exists regarding whether maspin acts as a protease inhibitor among biochemists.2022 Evidence from my laboratory indicates that maspin, regardless of whether it inhibits protease or not, possesses other functions independent of anti-protease action (Zhang et al, unpublished data).16
In this Chapter I intend to summarize the studies on maspin done by my group in the last few years and some works that are done by others but are closely related to ours. They will be divided into three sections: 1) maspin gene expression; 2) role of maspin in normal mouse development and 3) role of maspin in tumor progression and angiogenesis.
Maspin Gene Expression
Maspin Gene and Chromosome Localization
Human maspin encodes a 375 amino acid protein with a molecular weight of 42 K dalton. By FISH in situ analysis maspin is located at chromosome 18q21.3.13 To determine maspin gene structure, we have cloned maspin genomic DNA and sequenced all exon/intron borders as well as partial introns. Our data show that maspin gene contains 7 exons and 6 introns with a total length of 28 kb (Zhang et al, unpublished). It contains a non-coding exon 1 and an unusually long intron 1 (7.4 kb). Further genetic study and computer search of recently published human genome sequence shows that this region has a cluster of serpins belonging to the ovualbumin serpin family (Clan B). These include from the direction of telomere to centromere: PI8, PI10, PAI-2, megsin, SCCA1, SCCA2, headpin, and maspin.23,24 The gene structure of maspin is different from that of PAI2, SCCA1 and 2 in the same cluster since they all contain 8 exons and 7 introns. In addition, another cluster of serpins within the Clan B family exists in chromosome 6p25. Evidence from genetic study shows that these two clusters may have evolved from both intra- and inter-chromosome duplication.25
The phylogenetic tree shows that maspin is conserved with the ovalbumin family of serpins. The degree of homology at the amino acid level is : 40%, human monocyte-neutrophil elastase inhibitor (ELANH2), 39%, human PI10, 39%, Human PI8, 38%, human PI6, and 35% for human PAI-2.23,2628 ELANH2 and PI6 are located at chromosome 6p25.29 Other serpins at 18q21.3 such as PI8 are also more homologous to PI9 at 6p25, supporting the hypothesis of interchromosome duplication.
Differential Expression of Maspin in Breast Normal and Tumor Cells
The maspin gene was originally isolated from normal mammary epithelial cells. Northern blot analysis had been used to evaluate the expression pattern of maspin in human cell lines. Maspin was found to be highly expressed in 70N and 76N normal mammary epithelial cells, down-regulated in 21NT and 21PT primary breast tumors, and silent in a series of metastatic tumor cells. The gene was not expressed in cells of non-epithelial origin, such as mammary fibroblast cells, foreskin fibroblast cells, and U937 human monocytic cells. Mouse homolog of maspin was also expressed highly in mouse normal mammary epithelial cells such as NMuMG, HC11. The level of expression was down-regulated in low invasive mouse mammary tumor cells and lost in a highly invasive tumor cell line.
Transcriptional Regulation of Maspin in Breast Cells
To study maspin gene regulation in breast normal and cancer cells, we cloned maspin gene promoter. The cloning of maspin promoter has been quite a challenge because of the presence of a long intron 1. Most lambda genomic clones contain about a 20 kb genomic insert. Thus, using the full length of maspin cDNA as a probe, we were unable to recover any clones containing maspin promoter since the region from exon 2 to the last exon spanned over 20 kb. Finally, a YAC clone containing a cluster of serpins including maspin was used to screen maspin genomic DNA using a 50 mer oligomer as a probe. A positive clone was identified containing a 1.2 kb 5'-flanking region, a 7.4 kb intron 1 and partial exon 2 sequence. Sequencing and primer extension analysis showed that maspin did not have a typical TATA box in its promoter, a common feature for housekeeping or ubiquitously expressed genes. The major transcription start site was located 207 bp from first ATG site and the ATG was located in the second exon.17
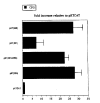
Several important transcription factor binding sites, Ets, Ap1, HRE and p53, are located within the 1 kb promoter region. To identify the responsive elements, promoter deletion and CAT assay were used to identify the activity of maspin in normal mammary epithelial cells (70N), primary tumor cells (21NT), and metastatic tumor cells (MDA-MB231) (Fig.1). As shown in Fig.1, a 1 kb upstream region was sufficient for activating transcription of maspin in normal breast cells. The activity was decreased in 21NT cells, and no detectable CAT activity was found in MDA-MB231 cells. Deletion analysis showed that deletion of an Ets site located at the region from −112 bp to −90 bp, abolished completely maspin activity, suggesting that this Ets was the major positivecis element within 1 kb responsible for up-regulation of maspin in normal mammary epithelial cells. Further analysis confirmed that this Ets site cooperated with a downstream Ap1 site for synergistic activation in normal breast cells; however this cooperative transactivation between Ets and Ap1 was lost in primary breast tumor 21NT cells.

Maspin Expression in Prostate Cells
Prostate cancer is the most common cancer in men. The prostate gland depends on androgenic hormones for its growth and development, analogous to the role of mammary hormones in development and morphology changes in the mammary gland.30 The molecular events leading to the development of prostate cancer may be similar to those in breast cancer. Because maspin functions as a tumor suppressor in the mammary gland, we have examined whether maspin could play a similar tumor-suppressing role in the prostate, and more importantly, what is the mechanism underlying gene regulation of maspin in prostate cells.
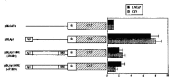
Northern blot analysis was carried out with RNAs from several human normal prostate and tumor cell lines. Maspin is highly expressed in CF3, CF91, and MLC normal prostate epithelial cells, and down-regulated in LNCaP, PC3, and DU145 prostate tumors. This expression pattern is similar to the findings in the normal mammary epithelial cells and carcinomas, indicating that the down-regulation of maspin expression is a common phenotype of both breast and prostate tumors.13,17 A 1 kb maspin promoter is active in CF3 normal prostate cells, but little activity is present in LNCaP prostate cancers. As in mammary epithelial cells, the same Ets site is primarily responsible for transcriptional action of maspin in CF3 cells. In addition, we have identified an HRE site located between −297 bp and −265 bp that represses transcriptional activity of maspin. Point mutations at this HRE site abolished the repression completely (see Fig. 2A.). The consensus HRE element has a sequence 5'-GGTACANNNTGT(T/C)CT-3'.31 This sequence can be recognized by multiple steroid receptors, such as glucocorticoid receptor, androgen receptor, and progesterone receptor.32 Previous studies have demonstrated that steroid receptors binding to HRE could mediate both transcription activation and repression.33,34 The HRE site (5'-GTACTCTGATCTCC-3') in the maspin promoter is unique in that its sequence does not share very good homology with the consensus sequence. This maspin HRE sequence acted as a general transcription repressor when it was placed in front of other heterologous promoters. Furthermore, we demonstrated that repression was mediated through AR using electrophoresis mobility shift assay (Fig. 2B). Binding of AR to maspin HRE is specific and can be blocked by AR antibody.35 These data may explain castration-mediated suppression of prostate cancer progression since ablation of androgen by castration may up-regulate maspin in prostate. From the therapeutic point of view, re-expression of maspin in the prostate tumors offers great hope for reversing the tumor phenotypes. Re-expression may be achieved by targeting both activation and repression modes. While it will likely be difficult to restore transcriptional activation of maspin through the Ets site, it may be more feasible to block the repression mediated by the AR binding HRE element. Treating tumors with ligands that block the binding of AR to the HRE or with reagents that compete strongly for binding to AR are possible methods of blocking HRE mediated repression. Our discovery of HRE-mediated repression offers another opportunity to increase the expression of maspin in prostate tumors, which may in turn reduce the progressiveness of prostate cancer.
Regulation of Maspin by Other Transcription Factors and Reagents
A number of groups have contributed to the discovery of new transcription factors and reagents that regulate maspin expression. Notably, a recent report has demonstrated that p53 activates maspin in prostate and breast cancer cells.36 Infection of these cancer cells through adenovirus p53 dramatically induced maspin expression with a rapid kinetics, similar to that of another well-known p53 regulated gene, p21. Inducers of p53 such as DNA-damaging reagents and drugs also induced maspin expression. They further showed that a consensus sequence for p53 binding is located in the maspin promoter and p53 binds to this site in gel shifting assay. A promoter luciferase reporter containing the p53 site was active when cotransfected with wild-type p53 into the prostate cancer cells. This finding is significant since it marks maspin as the first p53 regulated gene involved in cancer invasion and metastasis.
PPAR-γ plays an important role in adipocyte differentiation. A recent study also demonstrated that activation of PPAR-γ by its ligands caused dramatic morphological and nuclear changes that were characteristic of a more differentiated, less malignant state in breast cancer cells. Associated with these changes were the increase of maspin expression and arrest of cell cycle.37 Thus, PPAR-γ-mediated terminal differentiation of cancer cells might result from the increased maspin expression by a mechanism of transcriptional up-regulation. Gamma linolenic acid, an essential fatty acid, has anticancer properties. It was reported that gamma linoneic acid could induce maspin expression very rapidly.38 Cancer cells treated with GLA were shown to have reduced spreading and migration as monitored by video-microscope, and this reduction was reversed if the cells were treated with anti-maspin antibody. Thus, the anticancer effect of GLA was thought to result from the activation of maspin expression. Similarly, maspin was found to be induced by superoxide dismutase (MnSOD), and this induction might explain the tumor-suppressive effect of MnSOD in breast cancer cells.39
While it is relatively easy to identify the transcription factors and their regulatory sites responsible for maspin expression in normal cells, elucidating why certain cancers lose maspin may prove to be a more challenging task. Invasive cancer cells undergo numerous genetic changes such as mutation and chromosome rearrangement; the mechanism for loss of maspin expression in these cells is more complicated than just loss of a single transcription factor such as ETS. Indeed, one recent study has shown that in certain breast cancer cells, methylation of maspin promoter may contribute to the silencing of maspin in breast cancers.40
About ten-years ago, Dr. Ruth Sager proposed that tumor suppressor genes could be divided into two categories: class I and class II. Class I genes are mutated or deleted in tumors; examples include Rb and p53, which were identified by virtue of their mutations or deletions. Class II genes are not altered at the DNA level; rather, they affect the phenotype by changes in their expression levels.41 The focus of expression genetics is to re-express the class II tumor suppressor gene in cancers. Since the maspin gene is not mutated or deleted in tumors, it offers great advantage for one to induce maspin expression in tumors by the reagents identified from the above studies. In the future, some natural products or nontoxic drugs will likely be discovered to accomplish this goal.
Role of Maspin in Normal Development
From a genetic point of view, cancer is a developmental abnormality. To understand maspin's role in tumor progression, one needs to elucidate its normal function in organ development.
The mouse provides a model system for genetic manipulation. In comparison with human maspin, study of maspin in mouse has obvious advantages: 1) hormonal regulation of maspin is very difficult to study in human, while it is easy to obtain samples from mouse tissues such as mammary glands at different developmental stages; 2) transgenic animals can be produced to study the effect of overexpression of maspin on mammary gland development and tumor inhibition. Furthermore, knock-out mice can be produced to study the loss of function of maspin. For this purpose we cloned the mouse homolog of maspin.
Mouse Maspin is Highly Homologous to Human Maspin
Mouse maspin (mMaspin) cDNA was isolated from a mouse mammary gland cDNA library using human maspin cDNA as a probe. The longest cDNA sequence contained 1378 bp, including 69 bp 5' untranslated region, 1128 bp in the coding region, and 181 bp in the 3'-untranslated region. The poly (A) was added 23 bp after the polyadenylation signal site. The deduced amino acid sequence of mMaspin has a 89% identity with its human counterpart. In addition, a major difference in the mRNA lies in the 3' untranslated region. mMaspin cDNA has a short 3'-UTR, compared to the 1.2-kb 3'-UTR in the human maspin cDNA.16
Tissue Distribution
The maspin tissue expression pattern was examined by Northern blot analysis using RNAs from both human and mouse tissues. The human RNA blots from Clontech, Inc. contained a relatively small collection of tissues. Maspin was found to be abundantly expressed in prostate, thymus, testis, and small intestine.17 Among mouse tissues, the mammary gland has the highest level of expression. Maspin level varied during mammary gland development, suggesting its expression might be hormonally regulated. mMaspin is also expressed in mouse large intestine, skin, tongue, and stomach, and weakly in other tissues such as lung and thymus.16 It should be pointed out that Northern blot is not the ideal method to characterize maspin expression pattern. We have found out that some tissues express maspin at such a low level that it can not be detected by Northern blot analysis. One example is ovary, which has very low levels of maspin. However, deletion of one copy of maspin in heterozygous mice is enough to cause a severe defect in ovarian development (Zhang et al, unpublished data). Another feature noticed from Northern blot analysis was that there were multiple maspin transcripts in both human and mouse tissues.16,17 Alternative splicing may account for the multiplicity. Whether the splicing products play any biological role remains to be identified.
Functional Similarity Between Mouse and Human Maspin
Does mouse maspin function similarly as human maspin? To answer this question, we first compared the changes of maspin expression during mouse and human breast cancer progression.
Northern blot analysis showed that mouse maspin was expressed at a high level in mouse normal mammary epithelial cells (NMuMG, HC11). The level of expression was down-regulated in low invasive mouse mammary tumor cells (CSML0) and lost in a highly invasive tumor cell line (CSML100). The down-regulation of maspin protein was also confirmed by Western blot analysis. These data indicate that mouse maspin, like its human homolog, is also down-regulated during tumor progression.
Recombinant mMaspin proteins were produced in E. coli as a GST-maspin fusion protein. The biological function of recombinant protein was investigated using Boyden chamber assay (Fig. 3). A pair of mouse mammary tumor cells, CSML0 and CSML100 which are well characterized for their invasiveness, were used in both invasion and migration assays.42 Normally, about 10% of the low invasive CSML0 cells and 32% of the high invasive CSML100 cells seeded in the upper chamber invade the reconstituted basement membrane. Addition of mMaspin to the cell culture inhibits the invasion of both CSML0 and CSML100 cells. The inhibition by both cell lines increased as the concentration of GST-mMaspin was increased. The highest inhibitions at the concentration of 1.2 mM were about 60% for CSML0 cells (from 10% to 4%) and 62% for CSML100 cells (from 62% to 12.5%). For the motility assay, the recombinant proteins were added at different dosages to CSML0 and CSML100 cell suspensions and incubated with cells for 30 mins at room temperature before seeding. Under control condition, 5.8 % CSML0 cells and 7.2% CSML100 cells move through the polycarbonate membrane. Addition of GST-mMaspin inhibits the motility of both CSML0 and CSML100 cells. The highest inhibitions at the concentration of 1.2 μM were about 78% for CSML0 cells (from 5.9% to 1.25%) and 80% for CSML100 cells (from 7.2% to 1.4%).
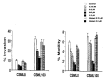
These data strongly suggest that mouse maspin functions like human maspin. Therefore, studying the function of maspin with transgenic and knock-out mice should provide valid insights into its role in the normal human breast and in breast cancers.
Transgenic Maspin Mice and the Role of Maspin in Normal Mammary Gland Development
Why Study the Role of Maspin in Normal Mammary Gland Development?
The mouse mammary gland undergoes a dramatic series of cyclical changes during development (see Fig. 4). In juvenile mice, the mammary gland develops based on a rapid, hormonally-regulated growth and morphogenesis of the epithelial ducts. Mammary ducts penetrate the gland by means of end buds. Upon reaching sexual maturity, the epithelial ducts have filled the whole gland with a tree-like structure. The onset of pregnancy initiates another stage of mammary development in which alveolar epithelial cells rapidly proliferate and differentiate under different regulatory mechanisms. Alveolar cells grow from the ductal skeleton and appear as lobulo-alveolar units by the end of pregnancy. These alveoli are the functional unit of milk production at lactation. Following lactation, the mammary gland undergoes massive remodeling and apoptosis, resulting in involution of the gland and a return to the ductal structure similar to the non-pregnant state.

Figure 4
Mouse normal mammary gland development. Adapted from the website: http://mammary.nih.gov/atlas.
One of the barriers to progress in breast cancer research is lack of the biological study of genes in normal mammary gland development. Since maspin expression is regulated during mammary gland development,43 it is absolutely relevant to examine the role of maspin in this process in order to better understand its involvement in breast cancer.
Establishing Maspin Transgenic Mice
To delineate maspin's function in vivo, we utilized a transgenic mouse system to examine the effect of overexpression of maspin under control of the whey acidic protein (WAP) promoter. The whey acidic protein has been shown to be exclusively expressed in mammary epithelial cells during midpregnancy and lactation.44,45 Transgenic mice were generated by injecting embryos with the WAP-maspin construct. Screening of founders was performed by Southern blot analysis using DNA isolated from mouse tissues. The expression of both endogenous maspin and the transgene in transgenic mouse mammary glands were examined qualitatively by reverse transcription-PCR. RNA was isolated from the mammary glands of transgenic mice at different stages of development. As expected, the WAP-maspin transgene was expressed during late pregnancy and lactation but was not detectable in virgin mice and glands undergoing involution.
Whole mount and histology of the mammary glands from wild-type and transgenic mice were analyzed. Ductal elongation and branching appeared to be normal in wild-type and transgenic virgin mice. No difference in alveolar structures were present between the transgenic and wild-type animals up to day 10 of pregnancy; however, minor differences did become noticeable at day 15 of pregnancy following the activation of the WAP promoter-driven transgene. These mammary glands exhibited decreased alveolar densities, which was further reduced as compared to controls at day 19 and resembled the morphology of the midpregnant wild-type controls. At high power, the mammary glands from transgenic mice contained not only fewer lobular-alveoli structures, but also the size of each alveolar structure was greatly reduced (Fig. 5). In many cases, the lumens of the alveoli were closed. This defect was due to the expression of the transgene activated by WAP promoter from midpregnancy.

Increased Apoptosis and Proliferation in Midpregnant and Early Lactating Mammary Glands of Transgenic Mice
Since the underdevelopment of the mammary glands in the WAP-maspin transgenic mice could have arisen from either decreased proliferation or increased apoptosis or a combination of the two, TUNEL and PCNA immunohistochemistry assays were carried out utilizing pregnant and early lactating mammary glands from wild-type and transgenic mice. As shown in Table 1, the apoptotic rate was significantly increased in transgenic glands at midpregnancy (2.29±0.26%) as compared to controls (0.91± 0.09%). In contrast, little difference was observed in cell proliferation at day 15 of pregnancy. However, during lactation, the apoptosis and proliferation profiles changed significantly in the transgenic strain. Secretive alveolar cells occupied the majority of the fat pad and there was a low rate of proliferation and apoptosis in samples taken from normal mammary glands. This observation contrasted with the results obtained from the WAP-maspin mice, in which a large percentage of the fat pad was devoid of alveolar cells and an increased rate of proliferation was observed. Both proliferation and apoptosis index decreased quickly as lactation proceeded, and by lactation day 10, very few cells were PCNA-positive and apoptotic in both transgenic and normal mice.
Table 1
The rate of apoptosis in wild-type and transgenic mice.
Effect of Transgene Expression on Milk Gene Expression
The defect in alveolar structures in the WAP-maspin mice during late pregnancy severely hampered the ability of the mother to successfully nurse her entire litter. Indeed, most of the pups died due to insufficient milk production. However, these pups could be rescued by fostering them to a BALB/c non-transgenic lactating female. The number of pups that a transgenic mother could nurse varied between animals. A survey of five sibling mothers at their first pregnancy yielded an average survival rate of 3.6 pups/litter.
Since milk protein genes can function as differentiation markers for the mammary gland, we compared their expression patterns in transgenic and wild-type control mice. Western blot analysis showed that WAP and β-casein were highly expressed in wild-type mammary glands at day 19 of pregnancy and during lactation. However, WAP and β-casein were not detectable in day 19 pregnant transgenic mice. Both milk proteins were present in lactating day 1 transgenic glands, but at a reduced levels, which increased as lactation progressed. This observed decrease was likely due to the effect of reduced number of alveolar cells and closed lumens in the late pregnancy transgenic mice.
Hypothesis for Maspin Function in Mammary Gland
We hypothesize that maspin may primarily regulate cell adhesion and motility in mammary cells, possibly by regulating integrin profiles. Indeed, expression of a dominant negative β1 integrin in the mammary gland, which disrupted the function of β1 and its associated integrins, resulted in a phenotype similar to that observed in the WAP-maspin transgenic mice.46 Both maspin and chimeric β1 transgene expression caused underdevelopment of the mammary gland in midpregnancy and early lactation, which was accompanied by an increase in apoptosis. In early lactation, milk protein levels were also reduced. The similarity in phenotypes suggests that overexpression of maspin may act to perturb integrin regulation or other associated cell adhesion molecules. This hypothesis is partially supported by an in vitro study, which showed that exogenous maspin modified cell attachment to fibronection by regulating integrin profiles, including down-regulation of β1 integrin.47
The mechanism by which maspin regulates apoptosis is also unknown. One possibility is that overexpression of maspin perturbs the adhesion of alveolar cells to the ECM as does the chimeric β1 transgene and, thus inhibits the motility of alveolar cells at a stage when invasion into the fat pad is critical. The proliferating alveolar cells are unable to migrate out, leading to increased apoptosis and a resulting small lumen. This is consistent with the concept that proper interactions of mammary epithelial cells with the basement membrane are essential for cell survival, and their disruption will trigger signals leading to apoptosis.48,49
Verification of these hypotheses requires identification of new maspin target proteins involved in cell adhesion and migration. We believe maspin possesses multiple functions and may use different domains for these functions (Zhang et al, unpublished data).18 Understanding the mechanism of maspin action will be greatly facilitated by the analysis of mammary phenotypes using maspin knock-out mice.
Maspin Knock-out Mice
A tumor suppressor by definition is a gene, when deleted or inactivated, would result in or facilitate the tumorigenesis of an organism. The two-hit hypothesis by Knudson suggests that loss of heterozygosity in a tumor suppressor gene will render the organism more susceptible to tumorigenesis. This prediction has been convincingly proved by numerous studies. For example, Donehower et al has demonstrated that p53 heterozygous mice can promote cancer formation and that loss of both p53 alleles is not a prerequisite.50 To prove maspin does function as a tumor suppressor, we applied gene disruption technique in mice to examine the effect of loss of the maspin function on mammary tumor progression, as well as other mouse development processes. Because of the difficulty to delete a large DNA fragment containing full length of gene, it is a conventional wisdom to target the deletion to a small region of genomic DNA. We have cloned mouse maspin genomic DNAs and designed a vector to selectively knock out the last exon encoding the RSL region and stop codon as well as the entire 3'-UTR. Since 3-UTR is responsible for mRNA stability, it is therefore unlikely that a truncated maspin protein can be made. To this date, we have already generated maspin knock-out mice by the approach described above with help from Dr. Philip Leder at Harvard Medical School and Dr. Francesco DeMayo at the Baylor College of Medicine.
The phenotype of the maspin knock-out mice is surprising. The homozygous mice are embryonic lethal. The existing knock-out mice for serpins and their protease targets are all viable. This demonstrates maspin indeed is a very unique and important serpin. Preliminary data by this laboratory indicate maspin is expressed in early stages of embryonic development (Zhang et al, unpublished data), and lethality demonstrates that maspin plays a critical role. Partial loss of maspin in heterozygotes also displays a phenotype. The heterozygous mothers are less likely to get pregnant and if they do, they deliver small litters of less than 4 pups, which can not be explained by the loss of homozygotes since wildtypes generally produce over 8 pups. Our data indicate that the ovulation efficiency of maspin heterozygotes is greatly reduced compared to control. Histology showed more than 50% of corpus luteum had entrapped oocytes along with hemorrhage in the center of corpus luteum, indicating ovary angiogenesis was affected. Using combined in vitro and in vivo approaches, we find out that maspin indeed plays an important role in the process of lutealization and atresia during ovarian development (Zhang et al, unpublished data). We are currently investigating the effect of loss of heterozygosity on mammary gland development and mammary tumor progression.
Role of Maspin in Tumor Progression and Angiogenesis
Mounting evidence has been collected about maspin function in tumor progression.51 In vitro cell culture studies demonstrated maspin inhibited tumor cell migration and invasion.15,16,52 Previously, many groups have reported the association of higher maspin level with better tumor prognosis.37,38,53 The key questions we would like to address are: 1) Does maspin inhibit tumor progression in immunocompetent mice? 2) How does maspin inhibit tumor progression?
Overexpression of Maspin on Tumor Progression in Bitransgenic Mice
To characterize maspin's in vivo activity during tumor progression, we generated transgenic maspin mice exhibiting targeted overexpression of maspin to mammary epithelial cells under the control of mammary specific whey acidic protein (WAP) promoter.43 Overexpression of maspin in normal mammary epithelial cells inhibits mammary gland development and induces apoptosis. Since the primary goal of generating WAP-maspin transgenic mice was to test the protective role of maspin overexpression on mammary tumor progression, we crossed WAP-maspin transgenic mice with a strain of oncogenic WAP-simian virus (SV) 40 T antigen (TAg) mice. WAP-TAg transgenic mice develop mammary tumors with 100% frequency and can be utilized to examine specific mechanisms of tumor progression at both early and late time points.54,55 The SV40 TAg initiates tumorigenesis through the inactivation of both p53 and the pRb related family of proteins.56,57 In human breast cancers inactivation of p53 function is found in up to 40% of tumors, and mutations in Rb or related proteins have also been reported.57,58
Overexpression of Maspin Reduced Tumor Growth and Vessel Density and Increased Apoptosis in Primary Adenocarcinomas
Both maspin overexpression and dominant gain of TAg expression induce mammary apoptosis, likely through different mechanisms. To determine if maspin overexpression could increase the level of TAg-induced apoptosis in mammary epithelial cells, the percentage of cells undergoing apoptosis in WAP-maspin/WAP-TAg double transgenic mice was compared to WAP-TAg single transgenic mice. Maspin overexpression increased the percentage of apoptotic cells from 1.8% in the single WAP-TAg transgenic mice to 5.6% in the WAP-maspin/WAP-TAg double transgenic mice (P< 0.01). In addition, bitransgenic mice exhibited a significant reduction in the number of alveolar-lobular structures and alveolar lumen size when compared to WAP-TAg single transgenic mice. Like WAP-TAg single transgenic mice, none of the WAP-maspin/WAP-TAg double transgenic mice could nurse their offspring beginning with the first litters. These results demonstrate that overexpression of maspin can increase the rate of apoptosis in preneoplastic mammary gland.
We then compared the rate of tumor growth of WAP-TAg and bitransgenic mice (Table 2). Tumors were measured biweekly after they were initially detected, and mice were euthanized when the primary tumor reached 2.5 cm in diameter. The tumor observation time (from the time of first palpable tumor to the time when the tumor reached 2.5 cm in diameter) was used to assess the rate of tumor growth. The presence of maspin overexpression significantly increased the time for a first palpable tumor to reach 2.5 cm in diameter from 36.3 to 49.3 days (p< 0.03).
Table 2
Analysis of primary and lung tumors.
To determine if maspin overexpression has an effect on microvessel density and apoptosis during tumor progression in vivo, microvessel density in adenocarcinomas with and without maspin overexpression were compared. Microvessel density was measured after CD31 staining. In small mammary tumors (tumor size £ 0.6 cm in diameter), the microvessel density was significantly reduced (p< 0.02), while the apoptotic index was significantly increased in the presence of maspin overexpression (p< 0.01).
Maspin Overexpression Decreased the Extent of Pulmonary Metastases but Did Not Affect Tumor Initiation
None of the TAg-/maspin+ and TAg-/maspin− mice developed any tumors after 1 year observation. In a separate experiment, 25 WAP-maspin transgenic mice (C57BL/6 background) were followed for more than 2 years, none of the mice developed spontaneous mammary tumors. However, both WAP-TAg and bitransgenic mice developed tumors at similar age. Within 24 weeks after they became pregnant, most of the WAP-TAg and bitransgenic mice developed mammary tumors. Thus, tumor initiation is TAg oncogene dependent.
To detect lung tumor metastasis, lung tissues were sectioned for microscopic analysis (Table 2). At the end point (all mice were sacrificed when primary tumor reached about 2.5 cm in diameter), bitransgenic mice did not show any difference from WAP-TAg mice in the number and weight of mammary tumors developed (data not shown). Serial sections were selected to score for micrometastatic tumor foci under high power microscope. Fifteen out of 26 (55.6%) WAP-TAg mice developed lung metastasis while in bitransgenic mice the rate of metastasis was greatly reduced to 37.5% (15 out of 40 bitransgenic mice). To compare the difference in the number of tumor foci of lung samples between these two species, the microscopic images of lung sections were captured, and the foci number/lung area was quantitated for each mouse. The bitransgenic mice had decreased foci numbers (0.356 /104 pixels) compared to that of WAP-TAg mice (0.655 /104 pixels). This information should be useful for future metastasis study.
Maspin as an Angiogenesis Inhibitor
Tumor metastasis is a multi-step process. Malignant tumors need to invade through the basement membrane, stromal tissues, and metastasize to other organs.59,60 However, for this process to occur, neovascular formation is required.61,62 This process, termed angiogenesis, provides tumors with nutrients and aids in the removal of metabolic wastes.63,64 The interaction between endothelial cells and tumor cells also generates a paracrine effect.65,66
To study the potential anti-angiogenic properties of maspin, the mouse maspin was produced inE. coli as a recombinant GST fusion protein and tested in a variety of angiogenesis assays. Recombinant maspin blocked endothelial cell migration induced by VEGF and bFGF in a dose-dependent manner with an ED50 of 0.2 μM-0.3 mM (Fig. 6). At 1 μM, maspin completely blocked the response of the endothelial cells to both angiogenic inducers, while the GST control was inactive. In vivo, purified maspin effectively inhibited neovascularization. Rat corneas were surgically implanted with non-inflammatory slow release pellets containing maspin with bFGF and examined six or seven days later for ingrowth of vessels. As shown in Fig. 7, maspin completely blocked bFGF-induced neovascularization. To determine if the anti-angiogenic activity of maspin depended on the inhibition of some undefined protease, several mutants were constructed, expressed and tested. The RSL (reactive serpin loop) near the C-terminus of serpin family members is essential for their anti-protease activity. Mutations at the RSL region of other serpins especially at the P1 site abolish serpin activity.67 To disrupt this loop in maspin two different mutations were introduced in the RSL region: a C-terminal deletion downstream of P7' residue16 (maspin DRSL) and a conversion of the critical P1 arginine of the RSL loop to a glutamine (maspin*). A third mutant was constructed in which the first 139 amino acids have been removed but the serpin region left intact (maspinDN). These defective mutants were tested on endothelial cells (Fig. 8). Those with RSL defects retained the ability to inhibit endothelial cell migration and mitogenesis. Protein with mutations in the RSL region also retained the ability to inhibit neovascularization in vivo. The N-terminal deletion, maspinDN, was defective in all assays so it was not possible to determine if a crucial active region had been deleted or if it was not a viable protein.
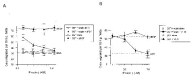

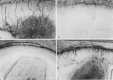
Complete inhibition of endothelial cell migration in vitro was achieved between 0.5 and 1 μM, in the same concentration range where maspin also inhibits tumor cell motility and invasion,15 but the mechanisms underlying these two maspin activities seem to be different. We and others find that the former requires that the protein have an intact RSL,16,52 whereas this feature was not essential for the inhibition of angiogenesis.
To determine if the ability of maspin to inhibit angiogenesis plays a role in its well-documented anti-tumor activity, an athymic mouse xenograft model was utilized. LNCaP prostate tumor cells were implanted subcutaneously on the bidorsal back of nude mice and tumor growth and neovascularization were monitored following systemic treatment with exogenous maspin. We found out that maspin-treated tumors contained significantly fewer vessels as determined by CD31 immunostaining than GST treated controls. To determine whether maspin effects on the tumor-induced vasculature were maintained during a more prolonged treatment, the above experiment was replicated with tumors harvested after 7 to 8 weeks. Thirty-two tumor sites were treated with maspin and 37 with GST. When examined at week 8, the growth of 53% of the maspin-treated tumors had been completely inhibited (Fig. 9). The remaining 15 maspin-treated tumors were reduced in size by average 3.43 fold when compared to GST control treated tumors. The effect of maspin was reversible. To examine if the reduced size of maspin-treated tumors coincided with reduced neovascularization, 20 representative tumors from either maspin-treated (10 sites) or GST-treated tumors (10 sites) were used to quantify the density of microvessels after immunostaining with CD31 antibody. The density of vessels in maspin-treated tumors was reduced 2.6 fold in average to that in control tumors and this difference was highly significant (see Fig. 9). We also compared the treated and control tumors of similar size. A reduction of vessel density was also observed in the maspin-treated samples.

The ability to inhibit tumor angiogenesis is only one of several activities associated with the intact maspin protein. Other serpins also have multiple functions and several of them are linked to angiogenesis and tumor growth.68 71 PAI-1 is involved in modulating both proteolysis and angiogenesis.12 PEDF, a known regulator of cell differentiation, is also a very potent anti-angiogenic factor.72 Such results indicate that a variety of molecules whose structure places them in the serpin family can be important regulators of natural tumor growth via their influence on neovascularization.
Future Directions
Since the discovery of maspin in 1994, a great deal of information has been generated regarding maspin's role in tumorigenesis, metastasis as well as normal development. More than 50 references have been published on maspin study. Despite these advancements, some critical questions remain to be addressed. What is the molecular mechanism of maspin action involved in anti-migration, tumor growth, and angiogenesis? How is maspin gene regulated during mammary gland development? Can we reexpress maspin in cancer cells to revert the phenotype by therapeutic intervention? I hope in the near future my laboratory, along with the colleagues in this field can successfully address these questions through a combined developmental and molecular approach. Understanding them is important for basic biology and will lead to therapies for cancer and other developmental diseases.
References
- 1.
- Worrall DM, Blacque OE, Barnes RC. The expanding superfamily of serpins: Searching for the real targets. Biochem Soc Trans. 1999;27(4):746–750. [PubMed: 10917676]
- 2.
- Potempa J, Korzus E, Travis J. The serpin superfamily of proteinase inhibitors: structure, function, and regulation. J Biol Chem. 1994;269(23):15957–15960. [PubMed: 8206889]
- 3.
- Whisstock JC, Irving JA, Bottomley SP, Pike RN, Lesk AM. Serpins in the Caenorhabditis elegans genome. Proteins. 1999;36(1):31–41. [PubMed: 10373004]
- 4.
- Tao W, Walke DW, Morgan JI. Oligomerized Ced-4 kills budding yeast through a caspase-independent mechanism. Biochem Biophys Res Commun. 1999;260(3):799–805. [PubMed: 10403845]
- 5.
- Wang YX, Turner PC, Ness TL, Moon KB, Schoeb TR, Moyer RW. The cowpox virus SPI-3 and myxoma virus SERP1 serpins are not functionally interchangeable despite their similar proteinase inhibition profiles in vitro. Virology. 2000;272(2):281–292. [PubMed: 10873771]
- 6.
- Hunt LT, Dayhoff MO. A surprising new protein superfamily containing ovalbumin, antithrombin- III, and alpha 1-proteinase inhibitor. Biochem Biophys Res Commun. 1980;95(2):864–871. [PubMed: 6968211]
- 7.
- Tombran-Tink J, Li A, Johnson MA, Johnson LV, Chader GJ. Neurotrophic activity of interphotoreceptor matrix on human Y79 retinoblastoma cells. J Comp Neurol. 1992;317(2):175–186. [PubMed: 1573062]
- 8.
- Brunner G, Preissner KT. Pericellular enzymatic hydrolysis: Implications for the regulation of cell proliferation in the vessel wall and the bone marrow. Blood Coagul Fibrinolysis. 1994;5(4):625–639. [PubMed: 7841321]
- 9.
- Blasi F. Urokinase and urokinase receptor: A paracrine/autocrine system regulating cell migration and invasiveness. Bioessays. 1993;15(2):105–111. [PubMed: 8385942]
- 10.
- Deng G, Curriden SA, Wang S, Rosenberg S, Loskutoff DJ. Is plasminogen activator inhibitor-1 the molecular switch that governs urokinase receptor-mediated cell adhesion and release? J Cell Biol. 1996;134(6):1563–1571. [PMC free article: PMC2120994] [PubMed: 8830783]
- 11.
- Stefansson S, Lawrence DA. The serpin PAI-1 inhibits cell migration by blocking integrin alpha V beta 3 binding to vitronectin [see comments] Nature. 1996;383(6599):441–443. [PubMed: 8837777]
- 12.
- Bajou K, Noel A, Gerard RD, Masson V, Brunner N, Holst-Hansen C. et al. Absence of host plasminogen activator inhibitor 1 prevents cancer invasion and vascularization. Nat Med. 1998;4(8):923–928. [PubMed: 9701244]
- 13.
- Zou Z, Anisowicz A, Hendrix MJ, Thor A, Neveu M, Sheng S. et al. Maspin, a serpin with tumor-suppressing activity in human mammary epithelial cells [see comments] Science. 1994;263(5146):526–529. [PubMed: 8290962]
- 14.
- Sun J, Rose JB, Bird P. Gene structure, chromosomal localization, and expression of the murine homologue of human proteinase inhibitor 6 (PI-6) suggests divergence of PI-6 from the ovalbumin serpins. J Biol Chem. 1995;270(27):16089–16096. [PubMed: 7608171]
- 15.
- Sheng S, Carey J, Seftor EA, Dias L, Hendrix MJ, Sager R. Maspin acts at the cell membrane to inhibit invasion and motility of mammary and prostatic cancer cells. Proc Natl Acad Sci U S A. 1996;93(21):11669–11674. [PMC free article: PMC38116] [PubMed: 8876194]
- 16.
- Zhang M, Sheng S, Maass N, Sager R. mMaspin: The mouse homolog of a human tumor suppressor gene inhibits mammary tumor invasion and motility. Mol Med. 1997;3(1):49–59. [PMC free article: PMC2230109] [PubMed: 9132279]
- 17.
- Zhang M, Maass N, Magit D, Sager R. Transactivation through Ets and Ap1 transcription sites determines the expression of the tumor-suppressing gene maspin. Cell Growth Differ. 1997;8(2):179–186. [PubMed: 9040939]
- 18.
- Zhang M, Volpert O, Shi YH, Bouck N. Maspin is an angiogenesis inhibitor. Nat Med. 2000;6(2):196–199. [PubMed: 10655109]
- 19.
- Sternlicht MD, Kedeshian P, Shao ZM, Safarians S, Barsky SH. The human myoepithelial cell is a natural tumor suppressor. Clin Cancer Res. 1997;3(11):1949–1958. [PubMed: 9815584]
- 20.
- Sheng S, Truong B, Fredrickson D, Wu R, Pardee AB, Sager R. Tissue-type plasminogen activator is a target of the tumor suppressor gene maspin. Proc Natl Acad Sci U S A. 1998;95(2):499–504. [PMC free article: PMC18448] [PubMed: 9435220]
- 21.
- Pemberton PA, Wong DT, Gibson HL, Kiefer MC, Fitzpatrick PA, Sager R. et al. The tumor suppressor maspin does not undergo the stressed to relaxed transition or inhibit trypsin-like serine proteases. Evidence that maspin is not a protease inhibitory serpin. J Biol Chem. 1995;270(26):15832–15837. [PubMed: 7797587]
- 22.
- Hopkins PC, Whisstock J. Function of maspin [letter; comment] Science. 1994;265(5180):1893–1894. [PubMed: 8091216]
- 23.
- Bartuski AJ, Kamachi Y, Schick C, Overhauser J, Silverman GA. Cytoplasmic antiproteinase 2 (PI8) and bomapin (PI10) map to the serpin cluster at 18q21.3. Genomics. 1997;43(3):321–328. [PubMed: 9268635]
- 24.
- Abts HF, Welss T, Mirmohammadsadegh A, Kohrer K, Michel G, Ruzicka T. Cloning and characterization of hurpin (protease inhibitor 13): A new skin-specific, UV-repressible serine proteinase inhibitor of the ovalbumin serpin family. J Mol Biol. 1999;293(1):29–39. [PubMed: 10512713]
- 25.
- Scott FL, Eyre HJ, Lioumi M, Ragoussis J, Irving JA, Sutherland GA. et al. Human ovalbumin serpin evolution: Phylogenic analysis, gene organization, and identification of new PI8-related genes suggest that two interchromosomal and several intrachromosomal duplications generated the gene clusters at 18q21-q23 and 6p25. Genomics. 1999;62(3):490–499. [PubMed: 10644448]
- 26.
- Schleef RR, Chuang TL. Protease inhibitor 10 inhibits tumor necrosis factor alpha-induced cell death. Evidence for the formation of intracellular high M(r) protease inhibitor 10-containing complexes. J Biol Chem. 2000;275(34):26385–26389. [PubMed: 10871600]
- 27.
- Sprecher CA, Morgenstern KA, Mathewes S, Dahlen JR, Schrader SK, Foster DC. et al. Molecular cloning, expression, and partial characterization of two novel members of the ovalbumin family of serine proteinase inhibitors. J Biol Chem. 1995;270(50):29854–29861. [PubMed: 8530382]
- 28.
- Harrop SJ, Jankova L, Coles M, Jardine D, Whittaker JS, Gould AR. et al. The crystal structure of plasminogen activator inhibitor 2 at 2.0 A resolution: implications for serpin function. Structure Fold Des. 1999;7(1):43–54. [PubMed: 10368272]
- 29.
- Sun J, Ooms L, Bird CH, Sutton VR, Trapani JA, Bird PI. A new family of 10 murine ovalbumin serpins includes two homologs of proteinase inhibitor 8 and two homologs of the granzyme B inhibitor (proteinase inhibitor 9). J Biol Chem. 1997;272(24):15434–15441. [PubMed: 9182575]
- 30.
- Schulze H, Isaacs JT, Coffey DS. A critical review of the concept of total androgen ablation in the treatment of prostate cancer. Prog Clin Biol Res. 1987;243A:1–19. [PubMed: 3309968]
- 31.
- Beato M. Gene regulation by steroid hormones. Cell. 1989;56(3):335–344. [PubMed: 2644044]
- 32.
- Laborda J. 36B4 cDNA used as an estradiol-independent mRNA control is the cDNA for human acidic ribosomal phosphoprotein PO. Nucleic Acids Res. 1991;19(14):3998. [PMC free article: PMC328497] [PubMed: 1861990]
- 33.
- Drouin J, Sun YL, Chamberland M, Gauthier Y, De Lean A, Nemer M. et al. Novel glucocorticoid receptor complex with DNA element of the hormone-repressed POMC gene. EMBO J. 1993;12(1):145–156. [PMC free article: PMC413186] [PubMed: 8428574]
- 34.
- Starr DB, Matsui W, Thomas JR, Yamamoto KR. Intracellular receptors use a common mechanism to interpret signaling information at response elements. Genes Dev. 1996;10(10):1271–1283. [PubMed: 8675013]
- 35.
- Zhang M, Magit D, Sager R. Expression of maspin in prostate cells is regulated by a positive ets element and a negative hormonal responsive element site recognized by androgen receptor. Proc Natl Acad Sci U S A. 1997;94(11):5673–5678. [PMC free article: PMC20837] [PubMed: 9159131]
- 36.
- Zou Z, Gao C, Nagaich AK, Connell T, Saito S, Moul JW. et al. p53 regulates the expression of the tumor suppressor gene maspin. J Biol Chem. 2000;275(9):6051–6054. [PubMed: 10692390]
- 37.
- Mueller E, Sarraf P, Tontonoz P, Evans RM, Martin KJ, Zhang M. et al. Terminal differentiation of human breast cancer through PPAR gamma. Mol Cell. 1998;1(3):465–470. [PubMed: 9660931]
- 38.
- Jiang WG, Hiscox S, Horrobin DF, Bryce RP, Mansel RE. Gamma linolenic acid regulates expression of maspin and the motility of cancer cells. Biochem Biophys Res Commun. 1997;237(3):639–644. [PubMed: 9299418]
- 39.
- Li JJ, Colburn NH, Oberley LW. Maspin gene expression in tumor suppression induced by overexpressing manganese-containing superoxide dismutase cDNA in human breast cancer cells. Carcinogenesis. 1998;19(5):833–839. [PubMed: 9635871]
- 40.
- Domann FE, Rice JC, Hendrix MJ, Futscher BW. Epigenetic silencing of maspin gene expression in human breast cancers. Int J Cancer. 2000;85(6):805–810. [PubMed: 10709100]
- 41.
- Sager R. Tumor suppressor genes: The puzzle and the promise. Science. 1989;246(4936):1406–1412. [PubMed: 2574499]
- 42.
- Tulchinsky E, Ford HL, Kramerov D, Reshetnyak E, Grigorian M, Zain S. et al. Transcriptional analysis of the mts1 gene with specific reference to 5' flanking sequences. Proc Natl Acad Sci USA. 1992;89(19):9146–9150. [PMC free article: PMC50082] [PubMed: 1329089]
- 43.
- Zhang M, Magit D, Botteri F, Shi Y, He K, Li M. et al. Maspin plays an important role in mammary gland development. Dev Biol. 1999;215:278–287. [PubMed: 10545237]
- 44.
- Pittius CW, Sankaran L, Topper YJ, Hennighausen L. Comparison of the regulation of the whey acidic protein gene with that of a hybrid gene containing the whey acidic protein gene promoter in transgenic mice. Mol Endocrinol. 1988;2(11):1027–1032. [PubMed: 2464745]
- 45.
- Pittius CW, Hennighausen L, Lee E, Westphal H, Nicols E, Vitale J. et al. A milk protein gene promoter directs the expression of human tissue plasminogen activator cDNA to the mammary gland in transgenic mice. Proc Natl Acad Sci U S A. 1988;85(16):5874–5878. [PMC free article: PMC281867] [PubMed: 2842753]
- 46.
- Faraldo MM, Deugnier MA, Lukashev M, Thiery JP, Glukhova MA. Perturbation of beta1-integrin function alters the development of murine mammary gland. EMBO J. 1998;17(8):2139–2147. [PMC free article: PMC1170558] [PubMed: 9545227]
- 47.
- Seftor RE, Seftor EA, Sheng S, Pemberton PA, Sager R, Hendrix MJ. maspin suppresses the invasive phenotype of human breast carcinoma. Cancer Res. 1998;58(24):5681–5685. [PubMed: 9865722]
- 48.
- Frisch SM, Francis H. Disruption of epithelial cell-matrix interactions induces apoptosis. J Cell Biol. 1994;124(4):619–626. [PMC free article: PMC2119917] [PubMed: 8106557]
- 49.
- Frisch SM, Ruoslahti E. Integrins and anoikis. Curr Opin Cell Biol. 1997;9(5):701–706. [PubMed: 9330874]
- 50.
- Venkatachalam S, Shi YP, Jones SN, Vogel H, Bradley A, Pinkel D. et al. Retention of wild-type p53 in tumors from p53 heterozygous mice: reduction of p53 dosage can promote cancer formation. EMBO J. 1998;17(16):4657–4667. [PMC free article: PMC1170795] [PubMed: 9707425]
- 51.
- Hendrix MJ. De-mystifying the mechanism(s) of maspin [news] Nat Med. 2000;6(4):374–376. [PubMed: 10742136]
- 52.
- Sheng S, Pemberton PA, Sager R. Production, purification, and characterization of recombinant maspin proteins. J Biol Chem. 1994;269(49):30988–30993. [PubMed: 7983035]
- 53.
- Xia W, Lau YK, Hu MC, Li L, Johnston DA, Sheng S. et al. High tumoral maspin expression is associated with improved survival of patients with oral squamous cell carcinoma. Oncogene. 2000;19(20):2398–2403. [PubMed: 10828881]
- 54.
- Tzeng YJ, Guhl E, Graessmann M, Graessmann A. Breast cancer formation in transgenic animals induced by the whey acidic protein SV40 T antigen (WAP-SV-T) hybrid gene. Oncogene. 1993;8(7):1965–1971. [PubMed: 8390039]
- 55.
- Li M, Hu J, Heermeier K, Hennighausen L, Furth PA. Expression of a viral oncoprotein during mammary gland development alters cell fate and function: Induction of p53-independent apoptosis is followed by impaired milk protein production in surviving cells. Cell Growth Differ. 1996;7(1):3–11. [PubMed: 8788028]
- 56.
- Dyson N, Buchkovich K, Whyte P, Harlow E. The cellular 107K protein that binds to adenovirus E1A also associates with the large T antigens of SV40 and JC virus. Cell. 1989;58(2):249–255. [PubMed: 2546678]
- 57.
- Li M, Lewis B, Capuco AV, Laucirica R, Furth PA. WAP-TAg transgenic mice and the study of dysregulated cell survival, proliferation, and mutation during breast carcinogenesis. Oncogene. 2000;19(8):1010–1019. [PubMed: 10713684]
- 58.
- Lee EY, To H, Shew JY, Bookstein R, Scully P, Lee WH. Inactivation of the retinoblastoma susceptibility gene in human breast cancers. Science. 1988;241(4862):218–221. [PubMed: 3388033]
- 59.
- Stetler-Stevenson WG, Aznavoorian S, Liotta L A. Tumor cell interactions with the extracellular matrix during invasion and metastasis. Annu Rev Cell Biol. 1993;9:541–573. [PubMed: 8280471]
- 60.
- Mignatti P, Rifkin DB. Biology and biochemistry of proteinases in tumor invasion. Physiol Rev. 1993;73(1):161–195. [PubMed: 8419965]
- 61.
- Folkman J. How is blood vessel growth regulated in normal and neoplastic tissue? G.H.A. Clowes memorial Award lecture. Cancer Res. 1986;46(2):467–473. [PubMed: 2416426]
- 62.
- Folkman J. Successful treatment of an angiogenic disease [editorial] N Engl J Med. 1989;320(18):1211–1212. [PubMed: 2469017]
- 63.
- Folkman J. Tumor angiogenesis. Mol Basis Cancer. 1995:206–232.
- 64.
- Fidler IJ, Ellis LM. The implications of angiogenesis for the biology and therapy of cancer metastasis [comment] Cell. 1994;79(2):185–188. [PubMed: 7525076]
- 65.
- Jones A, Harris AL. New developments in angiogenesis: a major mechanism for tumor growth and target for therapy. Cancer J Sci Am. 1998;4(4):209–217. [PubMed: 9689977]
- 66.
- Ambs S, Bennett WP, Merriam WG, Ogunfusika MO, Oser SM, Khan MA. et al. Vascular endothelial growth factor and nitric oxide synthase expression in human lung cancer and the relation to p53. Br J Cancer. 1998;78(2):233–239. [PMC free article: PMC2062907] [PubMed: 9683299]
- 67.
- Stringer HA, Pannekoek H. The significance of fibrin binding by plasminogen activator inhibitor 1 for the mechanism of tissue-type plasminogen activator-mediated fibrinolysis. J Biol Chem. 1995;270(19):11205–11208. [PubMed: 7744752]
- 68.
- Carmeliet P, Schoonjans L, Kieckens L, Ream B, Degen J, Bronson R. et al. Physiological consequences of loss of plasminogen activator gene function in mice. Nature. 1994;368(6470):419–424. [PubMed: 8133887]
- 69.
- Koolwijk P, van Erck MG, de Vree WJ, Vermeer MA, Weich HA, Hanemaaijer R. et al. Cooperative effect of TNFalpha, bFGF, and VEGF on the formation of tubular structures of human microvascular endothelial cells in a fibrin matrix. Role of urokinase activity. J Cell Biol. 1996;132(6):1177–1188. [PMC free article: PMC2120755] [PubMed: 8601593]
- 70.
- Montesano R, Pepper MS, Mohle-Steinlein U, Risau W, Wagner EF, Orci L. Increased proteolytic activity is responsible for the aberrant morphogenetic behavior of endothelial cells expressing the middle T oncogene. Cell. 1990;62(3):435–445. [PubMed: 2379237]
- 71.
- Min HY, Doyle LV, Vitt CR, Zandonella CL, Stratton-Thomas JR, Shuman MA. et al. Urokinase receptor antagonists inhibit angiogenesis and primary tumor growth in syngeneic mice. Cancer Res. 1996;56(10):2428–2433. [PubMed: 8625323]
- 72.
- Dawson DW, Volpert OV, Gillis P, Crawford SE, Xu H, Benedict W. et al. Pigment epithelium-derived factor: A potent inhibitor of angiogenesis. Science. 1999;285(5425):245–248. [PubMed: 10398599]
- The Role of Maspin in Tumor Progression and Normal Development - Madame Curie Bi...The Role of Maspin in Tumor Progression and Normal Development - Madame Curie Bioscience Database
- Rapid Evolution of Sex-related Genes: Sexual Conflict or Sex-specific Adaptation...Rapid Evolution of Sex-related Genes: Sexual Conflict or Sex-specific Adaptations? - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...