

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

There was never a time on earth when only one kind, one species, existed. At least there is no evidence to that effect and a plausible extension of that simple thought would lead to the conclusion that the development of species was parallel rather than sequential. The old paradigm starts with a single origin and in order to bring nature into harmony with that assumption, they invented speciation, to designate the splitting of species into new, biologically isolated units. Obvious as species are, speciation has never been substantiated and all examples given in the literature are postulates inspired by the hypothesis. The paradigm says that if animals A and B have certain features in common they are derived from a common ancestor. Thus, the evidence available is merely a restatement of the parent hypothesis.
During a conference on evolution in Prague in the 70s when a session on speciation began to grind to a halt I was asked to resuscitate the fighting spirit of the group. I do not remember why the onus fell on me after the organizers had allowed just minimal exposure for my thoughts on the program. There I was facing this group, their eyelids half folded, limbs hanging from the arm rests when I heard my own voice suddenly and unconnectedly asking whether any one in this group had ever seen unequivocal evidence for speciation. In a session on speciation this was the stone of Jason. Several jumped up and gave examples of multiple forms of a species that were tautologies on close examination, i.e., the hypothesis says that two similar looking animals came from one stem and therefore there was a speciation event; nothing really made a point. Finally one member of the group attempted a coup de grace for my contentiousness. Two colonies of fruit flies, he said, had been bred apart for the equivalent of millions of human propagation cycles and when brought together they would no longer interbreed! This was the core of the Darwinian idea confirmed in a simple indisputable experiment. What was there to say? Both groups still looked like Drosophila melanogaster, true, but it might have been seen as the initial stage of speciation; there was nothing left but to concede. At that moment a voice from the ranks broke the momentary silence, announcing that these experiments had long been shown to have been flawed and yes, the two groups were interbreeding as if they had never been separated. The session ran overtime without producing a single example of speciation. The excitement continued into the dining hall, but the lesson was lost over excellent Pilsner.
Yet, species are real and I owe the reader an explanation as to how they might have come about in the new model, particularly since earlier I argued for uniformity in chemistry. And by what means would one find an explanation if not by the same fossils that Darwinians are using combined with a new philosophy.
Even in our sort of orthodox climate of evolutionary thoughts experience teaches that fossils that are never unequivocal on nearly anything are very clear in denying the Darwinian process of speciation through imperceptible changes. Quite intelligent people are overlooking this conclusion because the evidence for the Darwinian model will be discovered, they are made to believe, during the next field trip. Of course that is an irrefutable argument, but it is also a worthless one. After 150 years of fruitless search it is prudent to conclude that the intermediates of the kind postulated by Darwinians do not exist.
Before even looking at fossils the genomist notes that evolution of animals via intermediates would be incompatible per se with the existence of species. The “inconceivably” large number of intermediates1 that anchor the old theory would have eliminated all species distinction and produced a bio-continuum instead, which is not observed. Furthermore, the fossil record shows species retaining their identity for millions of years, and that observation certainly does not support the concept of fluid barriers between them. Peculiar as it sounds, the Darwinian hypothesis in its original form is incompatible with the existence of species. On the other hand, species defined as propagating units are the only natural division of the fauna and flora; all higher-order taxonomic divisions are a way of grouping species by similarity, which may have no biological meaning
Species became apparent in the Ediacaren, but the picture is even more persuasive in the middle Cambrian period where the first arthropods were appearing, dense and varied, within a few centimeters of the lower layers of the Burgess shale.2 There is a splendid display in the Smithsonian Museum of Natural History of innumerable animals that appeared in a very short time, but the consequence of this observation has not even penetrated to the core of paleontologists’ paradigm which calls for about 10 million years for each speciation event. The reader may imagine a miniscule arthropod which would have been the ancestor of all others and therefore the beginning of the metazoan era. We will ignore the question as to how it propagated and come back to this spot in the shallow Cambrian waters about 10 million years later. On the Darwinian timetable there would now be two different kinds of arthropods, the one seen before, which had propagated into larger forms, and the smaller newcomer. This would have been the first speciation and, as one now visits the Cambrian every 10 million years it is noticed that, by the time the first road signs to the Ordovician appear, only six speciation events (10 million years each) have happened, which means 32 species materialized during the Cambrian as shown in Fig. 1.
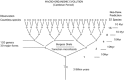
This conjecture collides with thirty major forms and 120 genera found in the bottom layer of the Burgess shale, which were deposited within the time span of a few million years. There they lie on top of each other like a cake of compressed creatures of the past.2 Adding to it the corresponding layer of the Cambrian Chengjiang fauna of China3 one cannot help but suspect that speciation had happened already long before the Cambrian or Ediacaren, although there are no hard fossils to confirm that suspicion. I have made a point of this message to my audiences and have noticed that it registered here and there.4–6
Of course, there are the calcareous microfossils of the Tomotian7 and the Precambrian,8 but nothing that would be an obvious immediate set of ancestors to the Cambrian assemblages. One knows that antecedents are there because of the laws of cause and effect, and we have even seen them in their pre-metamorphic phase without recognizing them because until now there was no reason to look for any such stages of life. The Genomic Potential Hypothesis should inspire paleontologists to search for instances of contiguous discontinuity (like in caterpillar and butterfly) in the layer that underlies the first fossiliferous horizon. This is a tall order, but recently the embryos of worms9 and possibly of arthropods10,11 have been discovered in the upper Precambrian layers. Microstructures, recovered from deep phosphate-rich deposits in China and Siberia,12 appear just like the proforms of animals might have looked that antedated the outpouring of phenotypes in the Ediacaran and the Cambrian which were followed periodically by more or less intense isplays up to the late Quaternary when upright walkers appeared. This would suggest that the Cambrian animals did not come from the Ediacaren group but derived more likely from germinal forms similar to those that had given rise to the Ediacaren fauna except that the Cambrian pro-forms were a little more complex! The pro-forms for the Devonian creatures were even more complex and remained silent for another 200 million years. My readers can continue the idea now until the Neogen when their own pro-form began to break into phenotypes. And why would that be a more reasonable conjecture than speciation by branching in the old model? Because there is evidence for the pro-form/phenotype transition whereas there is none for branching!
In proper perspective, i.e., in relation to a 3-billion-year-long developmental period, the distance on the time scale between the eruption of trilobites versus hominids is remarkably short. It is much too short to build a human from an arthropod by gene duplication and mutation!
Figure 2 depicts the Darwinian model as a single line from a single origin that swings up sharply at the beginning of the Cambrian. The multiple origin lines of the Genomic Potential Hypothesis symbolize genomic development that lasted 3 billion years for each life form, ending in the metamorphic state (wavy line) and finally in the same creatures that the Darwinian model must develop, one from another, in the phenomenally short period of 500 million years. The phenotypes of the Quaternary (our period) are so different from the Cambrian creatures, it is argued by Darwinists, that only a long time of adaptation, fine-tuning so to speak, by natural selection would have converted them into all subsequent species. One must object to that proposition on the following grounds: As the Precambrian stem cells burst onto the animal scene they brought along from the single-cellular (or colony) stage their appendices, sensory and reproductive organs, their feeding machinery, and armor for protection. But since these features, save reproduction, are without meaning in single-cellular life from which the animals just emerged, their appendages could not have been adaptations to any need for defense that multi-cellular life forms developed purportedly to deal with each other. What kind of evolutionary pressure would have prepared single-cellular organisms for what lay one short step ahead of them once they transgressed the line to multi-cellularity? The well-articulated legs and claws were an expression of genomic configurations that produced chitenous phenotypes directly and in prodigious amounts in the Cambrian. It was the chitin age; bone was a little down the road. Naturally one would ask if claws can come about at this stage on the basis of molecular development rather than adaptation, could not an arm come about by the same route a little later?

During Darwin’s time, St. George Mivart called attention to the difficulty entailed in the concept of adaptation by selection but it took 150 years until the modern evolutionists noticed and then they merely invented another term called pre-aptation instead of tossing the hypothesis.13
Why should one consider adaptation as an explanation for the production of new forms when there is no molecular mechanism and no material evidence? Extinction is prominent in the fossil record but not adaptation14 which is a concept that is a non-linear extrapolation of physiological adaptation, such as the increased myoglobin levels observed in the muscles of diving mammals, hemoglobin in mountain tribes, and muscle mass in athletes. These permissible adaptations always involve an increase or decrease of what is already part of an organism. The development of new body-parts is an entirely different matter that would, were it possible, involve changes in homeotic genes that control segment activity and co-ordination.
Metamorphosis, the process whereby one creature comes from one genome in two or three distinct forms called egg, caterpillar, and butterfly, is a well-recognized process because it can be observed during every phase. It is a prime example for phenotypical development by molecular design without guidance from Darwinian principles.
The transition from a primary ovum to a phenotype is an inescapable conclusion when all circumstances are considered. Just as clear is the fact that between the DNA sequences and the carapace of the first arthropods bilateral and multi-lateral symmetry fell into place without help from the environment.15 Most animals are symmetric, but there are enough exceptions to make the point that this is not due to ‘evolutionary pressure’ but is rather a consequence of a molecular configuration at the genomic level.
So what is the gist of all these explanations? They serve to make room for a new interpretation of the fossil record. According to the new model speciation was a fait accompli when the first animals began to produce fossils. It happened even earlier, the true edge-condition for speciation must be placed at the Hadean-Precambrian transition and that means that it was a chemical and not a biological phenomenon.
There are no species (as in life) before the Hadean transition, but the root of the argument concerning the segregation of future life forms (species) actually begins when nuclear raw material formed nucleotide sequences which would determine the possible outcomes of the developmental period of a cell. This is the absolute bottom of the story, the time when the genomic potential develops. Before polymerization, nucleotides do not carry any information, once they are linked into chains they spell out an organism if exposed to the proper conditions, and they remember their own sequence because of the complementary type of self-replication as plus and minus strands. There is a true border condition between monomers that carry information concerning chemical reactivity only and the polymers that hold the information for a macro-organism. For each future clone that consists of thousands of nearly alike nucleotide sequences in an area of biogenesis we can write the equation of species:
(Nucleotides 1, 2, 3, ---n) — polynucleotides-potential species + variants and so on for a vast number of biogenic foci.
The history of species begins in pools which were likely not much larger then a liter and which might have seen periods of concentration and dilution and what else the relatively rough climate of early earth presented. At the nucleotide level there was no readily discernible difference between the pools but when polymerization began every pool was different from its neighbor and chains of different sequences arose even in the same pool. When primary chains reached a certain length hydrogen bonding between a poly-nucleotide strand and free nucleotides would promote synthesis of complementary strands which would give a certain identity to a focal point. Information potential would increase with increasing length of the primary strand much like the information expressed as 0 and 1 on a Turing tape would increase with length of the tape. All of the processing of potential information would occur by the reactions that are well-known to molecular biologists such as loop formation, scission, fusion, and insertions. Furthermore, assembly of functions from a huge reservoir of nucleic acid, produced by untargeted polymerization, requires that large amounts of useless sequences exist. From what is known about the human genome that means 99% of the DNA in the nucleus.
Another interesting conclusion would be that such an assembly of untargeted information would contain a great deal of redundancy of important and of unimportant proteins. There are about 40 fibroblast growth factor genes of which the major forms can be experimentally eliminated (gene knockout) without any ill effects on the organism. A major blood protein in humans (albumin) is not required for survival or well-being, and relaxin appears in humans in two copies both of which are not essential. The pan-selection concept of the old model does not permit the production of useless products and modern gene technology proves them wrong almost daily by showing up unessential proteins. Clearly the organism is not constructed by reason and the remaining possibility is the assembly via an excess of potential functions. That observation leads us back to the pre-biotic origin of the only natural divisions of the fauna and flora, the species, and there is really no viable alternative, nothing that could stand up to the demands of epistemology. This line of reasoning leads to a universal concept of biogenesis and bio-diversity, which states that the complexity and diversity of life on a planet is a function of the amount of nucleic acids that have accumulated via chemical synthesis.
References
- 1.
- Darwin C. The Origin of Species by Means of Natural Selection New York: The Modern Library of New York, 1859.
- 2.
- Morris SC. The Cambrian “explosion”: Slow-fuse or megatonnage? Proc Natl Acad Sci USA. 2000;97:4426. [PMC free article: PMC34314] [PubMed: 10781036]
- 3.
- Chen J -Y, Ramskoeld L, Zhou G -q. Evidence for monophyly and arthropod affinity of Cambrian giant predators. Science. 1994;264:1303. [PubMed: 17780848]
- 4.
- Ohno S. The notion of the Cambrian pananimalia genome. Proc Natl Acad Sci USA. 1996;93:8475. [PMC free article: PMC38696] [PubMed: 8710894]
- 5.
- Doolittle WF. Phylogenetic classification and the universal tree. Science. 1999;284:2124. [PubMed: 10381871]
- 6.
- Woese C. The universal ancestor. Proc Natl Acad Sci USA. 1998;95:6854. [PMC free article: PMC22660] [PubMed: 9618502]
- 7.
- Stanley SM. Earth and Life Through Time Viking Press,New York, 1986.
- 8.
- Schopf JW. Solution to Darwin's dilemma: Discovery of the missing Precambrian record of life. Proc Natl Acad Sci USA. 2000;97:6947. [PMC free article: PMC34368] [PubMed: 10860955]
- 9.
- Chen J -Y, Oliveri P, Li C -W. et al. Precambrian animal diversity: Putative phosphatized embryos from Doushantuo Formation of China. Proc Natl Acad Sci USA. 2000;97:4457. [PMC free article: PMC18256] [PubMed: 10781044]
- 10.
- Pratt BR, Zhang X -G. Unscrambling fossilized Cambrian embryos. J NIH Res. 1995;7:46.
- 11.
- Bengtson S, Zhao Y. Fossilized Metazoan embryos from the Earliest Cambrian. Science. 1997;277:1645.
- 12.
- Li C -W, Chen Y -J, Hua T -E. Precambrian sponges with cellular structures. Science. 272;279:879. [PubMed: 9452391]
- 13.
- Lewin R. Adaptation can be a problem for evolutionists. Science. 1982;216:1212. [PubMed: 17830574]
- 14.
- Raup DM. Biological extinction in earth history. Science. 1986;231:1528. [PubMed: 11542058]
- 15.
- Peterson KJ, Davidson EH. Regulatory evolution and the origin of the bilaterians. Proc Natl Acad Sci USA. 2000;97:4430. [PMC free article: PMC34315] [PubMed: 10781037]
- The Origins of Species - Madame Curie Bioscience DatabaseThe Origins of Species - Madame Curie Bioscience Database
- Telomerase Activity as a Marker of Tumor Cell Survival to Evaluate Sensitivity o...Telomerase Activity as a Marker of Tumor Cell Survival to Evaluate Sensitivity of Neoplastic Cells to Cancer Treatment - Madame Curie Bioscience Database
- Nuclear Envelope Dynamics in Drosophila Pronuclear Formation and in Embryos - Ma...Nuclear Envelope Dynamics in Drosophila Pronuclear Formation and in Embryos - Madame Curie Bioscience Database
- Cytokines in the Pathogenesis of Rheumatoid Arthritis and Collagen-Induced Arthr...Cytokines in the Pathogenesis of Rheumatoid Arthritis and Collagen-Induced Arthritis - Madame Curie Bioscience Database
- Evolutionary De Novo Design - Madame Curie Bioscience DatabaseEvolutionary De Novo Design - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...