

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Introduction
The temporal organization of Euglena gracilis Klebs (strain Z) has been studied extensively over the past 40 years (reviewed in Refs. 1-5). This algal flagellate can be grown on a variety of defined media, either photoautotrophically in the presence of CO2 and vitamins B1 and B12, or organotrophically in the light or dark on carbon sources ranging from acetate and ethanol to lactic, glycolic, glutamic and malic acids over a wide pH range. This versatility in growth mode, in conjunction with the fact that cell division can easily be synchronized by 24-h (and other) lighting schedules6,7 and temperature cycles,8 has made Euglena an important experimental organism for physiological and biochemical investigations.9-11
A number of persisting circadian rhythms (CRs) have been reported (Table 1). These studies have been aided by the fact that Euglena can be maintained in the “stationary” growth phase (infradian growth mode) for days or weeks with little or no net change in cell titer; circadian output can be monitored while divorced from the driving force of the cell division cycle (CDC). The fact that a number of photosynthetic mutants (even completely bleached strains devoid of their chloroplast genomes) have been isolated that still exhibit light-entrainable CRs effectively has eliminated the problem of the dual use of imposed light spans and signals—as an energy source for growth and as a timing cue for the underlying clock.12-23
The Euglena system thus provides an excellent case for temporal differentiation: diverse behavorial, physiological and biochemical activities are partitioned along the 24-hour axis, providing a dimension for environmental adaptation and functional integration in time.2,24 This circadian time structure is well illustrated by an acrophase chart (fig. 1), which provides a convenient way to indicate relationships of rhythms (analyzed mathematically by cosinor or other techniques) both to a synchronizing LD cycle (or other Zeitgebers) and to each other during either entrainment or free-running conditions.3

Survey of Selected Classes of Circadian Rhythms in Strains
In the following subsections, we survey in greater detail selected CRs itemized in Table 1 that have been reported since the last major review2 and examine some of the more recent developments in these areas.
Table 1
Circadian time structure in Euglena gracilis Klebs.
Cell Division Rhythmicity in Wild-Type and Mutant Euglena
The CDCs of many algae, fungi and protozoans exhibit persisting CRs of cell division, or “hatching”.24 Division occurs only at a certain phase of the circadian cycle—often times (“subjective” nights) in constant darkness (DD) or light (LL) corresponding to the dark intervals in a synchronizing LD cycle. This “gating” phenomenon, reflecting an interaction between a circadian oscillator (CO) and the CDC, has been intensively investigated in Euglena.2 Because it will serve as a basis for the analysis of the mechanism whereby coupling between the clock and cell cycle is achieved, we treat this class of rhythm in depth.
Wild-Type Strain
The canonical properties of circadian clocks—entrainment by diurnal and non24-h light6,7,26-28,81 and temperature8,55 cycles, persistence,82 initiation, phase-shiftability,83 phase singularity84 and temperature compensation85—have been found to characterize the rhythm of cell division in the photoautotrophically grown, green wild-type strain. The latter two properties deserve further mention.
Singularity Point
Mathematical studies have predicted that a CO might be rendered arrhythmic—characterized by a phaseless, motionless state—by a critical pulse of a certain strength and duration given at a specific time (termed the “singularity point,” S*) in the circadian cycle.86,87 As stimulus strength is increased, the transition from Type 1 (weak pulse) to Type 0 (strong pulse) resetting is necessarily discontinuous at one special phase point, the “breakpoint,” corresponding to this unique singularity. This prediction hasbeen demonstrated for light and chemical perturbations in several circadian systems,88 and the clock underlying the cell division rhythm is no exception.
The threshold intensity of illumination for phase-shifting the division rhythm in Euglena by 3-h light signals has been examined.84 Perturbations having an intensity of 700 to 7500 lux generated the same phase shift (Δ℘) at a given CT (fig. 2A). Signals of lower intensity, however, elicited different responses. Thus, a 40- to 400-lux pulse given at CT0.4 (the approximate location of the breakpoint at about CT23) induced arrhythmicity (fig. 2B), the population reverting to asychronous, exponential growth. The intensity of this annihilating pulse and the CT at which it was imposed were quite specific: A 300-lux stimulus given at CT21.5 (fig. 2C) merely generated a phase delay (-Δ℘) of the same magnitude found for 7500-lux signals. Different degrees of asynchrony were observed as one approached the boundaries (lux and CT) of the critical pulse. The location of S* at approximately CT24 (CT00) corresponds to the steepest part of the Type 0 resetting curve (see Ref. 83) and probably lies on the axis of the three-dimensional phase resetting surface of Euglena.
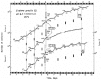
The existence of this “critical pulse” and its corresponding S* not only further supports the hypothesis that a CO regulates the CDC in Euglena but also suggests that the pacemaker may have limit cycle dynamics. Inasmuch as the results were obtained with populations of cells, we cannot deduce the state of the oscillator(s) in individual cells. Arrhythmicity in the culture may be the result of a dispersion of the phases of individual CDCs (cellular incoherence), entailing that the pulse differentially phase shift the cells which were synchronous at the time of the pulse; or, the critical pulse may have stopped each circadian clock comprising the population (but not the CDCs, which continue to run).
Temperature Compensation
A remarkable property of CRs is that their period, but not their amplitude, is only slightly affected by the ambient steady-state temperature over the physiological range.89 This is just what one would anticipate in a functional biological clock measuring the solar day. In contrast, the duration of the CDC is highly dependent on temperature, and, indeed, this is true for Euglena.8 A study of the effects of different constant temperatures ranging between 16 and 32°C on the generation time (g) of the wild-type Z strain (as well as the DCMU-resistant ZR line) photoautotrophically batch-cultured in LD:3,3 clearly illustrates this dependency (fig. 3A).85

In a more extensive comparative study of temperature compensation of the free-running period in the Z and ZR strains of Euglena maintained in LD: 3,3 at different steady-state temperatures,85 a Q10 of 1.05 was found in the former (fig. 3B), indicating that it was virtually unaffected by changes in temperature over a 10°C range (22 to 32°C). (The circadian clock was not as well compensated in ZR, in which an average value for the Q10 of 1.23 was observed over a temperature range of 18 to 28°C. Nevertheless, even the CO of ZR was affected only modestly by temperature as compared to typical biochemical reaction rates and other biological processes, such as membrane transport.) Finally, a critical permissive temperature of 22°C was found for the Z strain (18°C for ZR): Although slow exponential growth occurred at lower temperatures, synchrony was not seen.)
Photoinduced Commitment and Photoperiodic Control
Earlier work with Euglena and the green alga Chlamydomonas90-94 has suggested that the CDC comprises two consecutive processes: a precommitment stage and a commitment stage. Progress through the former during the G1 phase of the CDC would be dependent on photosynthetic growth. The A point denotes the time at which the progression of G1-phase cells is light-dependent, whereas point T marks the time when cells become able to (i.e., are commited to) complete the CDC, even in darkness.90 If cells in the precommitment stage are transferred to DD, they immediately become arrested somewhere between the A and T points, whereas cells in the commitment stage, when transferred to DD, proceed through the remainder of the CDC until they stop at A in the following cycle. This photosynthetic regulation of CDC progression in algal cells resembles the nutritional control of heterotrophs. Nutritional deprivation in Saccharomyces cerevisiae or Schizosaccharomyces pombe arrests the CDC in either the G1 or G2 phase,95-97 while yeast cells that have progressed beyond “Start” (at the G1/S border) or a size-control checkpoint are committed to complete the CDC even if they are undergoing starvation.98
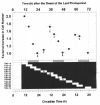
In a study of the effects of light and darkness on CDC progression during log-linear photoautotrophic growth of Euglena, Hagiwara et al30 demonstrated both dark-induced CDC arrest and photoinduced commitment to cell-cycle transition. Light-dependent restriction points were found, not only in the G1 (as previously described for Chlamydomonas), but also in the post-G1 phases as well. Thus, if cultures were transferred from LL to DD, some (committed) cells in the G1, S or G2 phases were able to undergo one or two CDC transitions even in DD, eventually becoming arrested in S, G2 or the subsequent G1 after mitosis. Other (uncommitted) cells were not able to proceed in the CDC. Committed cells that received light of higher intensity were more likely to undergo cell cycle transition in DD and to commit earlier at each CDC phase. Commitments were not achieved at fixed phase-points in the G1, S or G2 phases of the CDC.
The commitment to CDC transit in Euglena, therefore, is regulated by the maturity of the cell-cycle phases that is comprises and by light intensity. In addition, Hagiwara et al31 have demonstrated that a CO regulates these light responses of commitment to CDC phase transitions. When algal cells were entrained by LD:14,10 and then transferred to DD at the eighth hour of the last LD photoperiod, cells were arrested in G1, S or G2. Subsequent exposure of these dark-arrested cells to a 6-h light pulse at different CTs allowed them to further proceed in the CDC in a manner that was dependent on the CT of the pulse (fig. 4). Maximum photoinduction occurred around dusk (CT12). These results demonstrate a circadian gating of photoinduction of commitment to cell-cycle transitions—not just of cell division, as discused earler in this section—and the photoperiodic regulation of cell reproduction. Results from inhibitor experiments with diuron (DCMU), 2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone (DBMIB), methyl viologen, N,N,N',N-tetramethylphenylenediamine (TMPD) and carbonyl cyanide-m-chlorophenylhydrazone (CCCP) suggested that the photoinduced commitment of G2 cells to cell division required light for a signal originating upstream in noncyclic photosynthetic electron transport, particularly cytochrome b6-f, but not for the metabolic energy required for the process.31
Mutant Strains
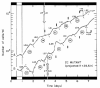
In order to eliminate a cellular compartment in which the clock might lie and to obviate possible signaling influences of the LD cycles previously used to elicit rhythmicity and provide energy for growth, we have chosen the achlorophyllous ZC mutant of Euglena—an obligate organotroph—for a series of studies parallel to those carried out in the wild-type strain.16 Division rhythmicity could be entrained by LD:12,12, or even by a one-pulse skeleton photoperiod (LD:1,23), and the rhythm free-ran in ensuing DD for at least 8 days (τ = 25.5 h), providing that the overall growth rate, or generation time (g) was >24 h (see fig. 5). Similar, though less extensive results had been obtained earlier with other photosynthesis mutants of Euglena, such as the white, heat-bleached, achlorophyllous W6ZHL strain and the ultraviolet light-induced P4ZUL mutant.1,13,15,26 A Type 0 phase-response curve (PRC) for light signals in the ZC mutant strain was generated from 15 phase-shifting experiments16 (fig. 5); this PRC (fig. 6) resembled that previously obtained with the wild-type strain.83 These studies have provided even more conclusive evidence for the role of a CO in the control of the CDC and have effectively circumvented the problem of the dual use of imposed LD cycles: as an energy source, or “substrate,” for growth and as a timing cue (Zeitgeber) for the underlying clock.
Thus, the CO plays a key role in the control of the CDC in Euglena, taken to be representative of other eukaryotic cells. Mitosis would not be an essential part of the oscillator but would lie downstream from it. Blockage of cell division should not stop the system from oscillating. 29,99 Therefore, cell division would be a “hand” of the underlying clock. We have tested this hypothesis in the wild type in two ways: (i) If the free-running division rhythm was stopped due to low initial levels of vitamin B12, and if this inhibition subsequently was released by readdition of B12 to the medium, the rhythm started up again in phase with an unperturbed control. (ii) If a pulse of lactate was given to a free-running culture, temporarily accelerating the CDC and overriding the CO,13 the phase of the rhythm when it was finally restored after the supplemental substrate had been depleted was in phase with that of an unperturbed control. These results are consistent with those found previously for the in-phase restoration of rhythmicity in the P4ZUL mutant free-running in LL by addition of sulfur-containing compounds to the medium.15
Respiratory Activity in Colorless Euglena Mutants
A CR of photosynthesis has been reported23 in cultures of wild-type E. gracilis Z cultures, in which the concentration of free O2 in the medium was continuously measured with a Clark electrode. Oxygen production peaked in the middle of the day, and the rhythm persisted in LL for a least 8 cycles, thus confirming earlier findings41,44 (see Table 1). These data were extended to colorless mutants lacking functional chloroplasts,23 but with this strain the phase of the rhythm was the inverse of that in the wild-type strain, with the highest values of oxygen concentration in the medium occurring during (subjective) night (fig. 7). The rhythm in the mutant cells responded directly to the light of more complex LD regimens, and red light (>598 nm) and blue light (<550 nm) were equally effective in regulating respiration activity as compared to white light if given as LD:8,8 (which synchronized the cells). Thus, the demand for energy, provided by either photosynthesis or respiration depending on the cell type, is under both light and circadian clock control. The origin of the rhythm in photosynthesis may lie in the light-induced synthesis of light-harvesting proteins (LHCPs) asssociated with the two photosystems, whose synthesis is clock-regulated, showing an up to 20-fold increase in the subjective day (see following section).76
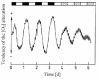
Circadian Synthesis of Light-Harvesting Proteins
By the in vivo labeling of proteins with [35S]methionine in cultures of E. gracilis grown in LL, Künne and de Groot64 confirmed earlier observations63 that total protein synthesis was circadian, and, further, that the τ was only slightly influenced by temperature. Three proteins (separated by SDS-PAGE), having molecular weights of 17, 24 and 60 kD, were found to be synthesized rhythmically (τ ∼26 h).76 The 60 kD protein had a maximal rate of synthesis at about CT11, while the two smaller proteins showed peaks at approximately subjective noon (CT05–08) (fig. 8). Their rate of synthesis oscillated by up to 20-fold when maxima and minima were compared. Over temperatures ranging between 16 and 27°C, τ differed by only 3 to 5 h, reflecting fairly good temperature compensation of the free-running period. Synthesis of the two smaller polypeptides was inhibited by cycloheximide but not by chloramphenicol, implying that synthesis occurred on 80S ribosomes. Once synthesized and processed, they were fairly stable in LL for about 120 h, although they were slowly degraded in darkness.
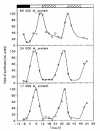
Subsequently, the two smaller proteins were identified as belonging to the family of light-harvesting-chlorophyll-proteins (LHCPs) of class I (17 kD) and class II (24 kD) by N-terminal sequencing of proteins isolated by two-dimensional PAGE and by detection with an anti-LHCP II serum.77 Although the total amount of LHCPs remained almost constant in LD:12,12, the amount of newly synthesized 17 and 24 kD proteins varied by about 20-fold, with maximum synthesis in the light phase. In contrast, their specific mRNAs varied only slightly, a finding (confirmed by using inhibitors of transcription) that suggests that the circadian control of LHCP synthesis occurs at the translational level, in contrast to most higher plants. It may well be that these rhythmicities in the synthesis of LHCPs play a significant role in the circadian control of photosynthesis in Euglena (see Table 1).
Circadian Rhythms in Enzymatic Activity
The activity of a number of enzymes have been shown to undergo circadian variation in Euglena (Table 1). Some more recent representative examples follow.
Aromatic Amino Acid Metabolism
Two key enzymes of aromatic amino acid metabolism, tryptophan 2,3-dioxygenase (TDO) and tyrosine aminotransferase (TAT) exhibit different rhythmic patterns in E. gracilis Z.69 These two enzymes are interrelated by common regulatory mechanisms, especially with regard to the control of gluconeogenesis from amino acids. Whereas TDO displayed a pronounced circadian rhythm in LL (peak activity at ∼CT18), TAT oscillated with an ultradian periodicity (τ = 4.26 h). Even in LD:12,12, TAT exhibited an ultradian period (∼4.63 h). From a dynamical viewpoint, therefore, these two enzymes can be dissociated. Under the same conditions, other variables such as phototaxis, cell shape and total protein synthesis also are under circadian control (see Table 1).54
NAD Kinase, NADP Phosphatase and the Mitochondrial Calcium Cycle
The in vivo levels of NAD+, NADP+ and NADPH were measured in synchronously dividing and in very slowly dividing cultures of E. gracilis Z grown photoautotrophically in LD:3,3 and were found to oscillate (τ = 27 h), with an amplitude not correlated with the CDC (fig. 9).68 There was also a CR in the activity of NAD+ kinase (peak at CT00) in extracts of Euglena with a phase relationship such that it could induce the rhythm in the in vivo level of NAD+ (see Goto100). No circadian oscillation of the ratios of either NADH/(NAD+ + NADH) or NADPH/(NADP+ + NADPH) occurred. Finally, a 3-h light pulse applied at CT18 to the free-running division rhythm not only generated the expected Δ℘, but also similarly reset the phase of the oscillation in total NAD+.68 These results suggest that the circadian oscillation in the in vivo level of NAD+ could be ascribable, as for the duckweed, Lemna,100 to that of the conversion between NAD+ and NADP+, but not to that of reduction–oxidation between NAD+ and NADH.
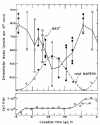
To determine whether NAD+ constitutes a possible clock “gear,” small seed cultures displaying a free-running circadian rhythm of cell division were given 2-h pulses at various CTs with NAD+ or NADP+—expected to directly elevate their own in vivo levels and then to increase or decrease the rate of the reactions catalyzed by NAD+ kinase and NADP+ phosphatase —or p-nitrophenylphosphate, a competitive inhibitor of NADP+, and then resuspended in fresh medium (effectively terminating the pulse) for subsequent monitoring of the division rhythm.68 PRCs were derived for each of these agents. Furthermore, a 2.5-h pulse of NAD+ given at CT21.7, which caused a Δ℘ of 4 h in the division rhythm, also phase-shifted the CR in the in vivo levels of of NAD+, NADH, NADP+, and NADPH. These results suggest that NAD+ (or NADP+, or NADPH), NAD+ kinase, and NADP+ phosphatase might represent components of the underlying CO.
What is the element that regulates these enzymes? Inasmuch as Ca2+-calmodulin activates NAD+ kinase in many plants, including green algae, and in the sea urchin, and because the CRs in the activities of NAD+ kinase and NADP+ phosphatase appear to be generated by a rhythm in the in vivo level of this complex in Lemna,100 it would be a likely candidate for a clock element in Euglena also. Attempts were made to perturb the clock either by directly causing a transitory decrease in [Ca2+] with 2- to 3-h pulses of chlortetracycline, a membrane-permeable chelator of Ca2+, or by transitorily inhibiting Ca2+–calmodulin by similar short pulses of W7 and chlorpromazine, both calmodulin inhibitors.68 All three drugs phase-shifted the division rhythm. Their PRCs suggested that both cytosolic Ca2+ and calmodulin constitute clock gears. If so, there should be another gear directly regulating the level of Ca2+ in Euglena. The main regulatory sites for many noncircadian systems are known to be the plasmalemma, the endoplasmic reticulum, and the mitochondria. To test the possibility that the mitochondrial Ca2+ -transport system might be a clock gear,68 cultures were pulsed (2 to 3 h) with either nitrogen, dinitrophenol, or sodium acetate, all of which affect electron transport, ATP hydrolysis and mitochondrial Ca2+-efflux. Each of these agents phase-shifted the division rhythm.
Based on these findings, Goto et al68 proposed a model in which NAD+, the mitochondrial Ca2+-transport system, Ca2+, calmodulin, NAD+ kinase and NADP+ phosphatase would represent clock elements that in ensemble, might constitute a self-sustained, circadian oscillating loop—not “the” clock but an oscillator in what is most probably a cellular ”clock-shop.” This proposed feedback loop can autonomously oscillate because the “cross-couplings” are always of opposite sign. The rhythmic activities of the two enzymes would then oscillate 180° out of phase because of the circadian changes in cytoplsmic concentration of one of their effectors, the Ca2+–calmodulin complex, which would activate NAD kinase but inhibit NADP phosphatase.
Subsequent in vitro studies9,20 detected circadian variations in the activities of NAD kinase and NADP phosphatase in the soluble and membrane-bound fractions of both synchronously dividing and nondividing cultures of the ZC mutant of Euglena in DD. Bimodal CRs in total NADP phosphatase activity were found in dividing cells (peaks at CT00 and CT12). The peak observed at CT00–03 disappeared when the cells had ceased dividing, a result that suggests that it might be regulated by the CDC. NAD kinase activity displayed unimodal CRs (peak at CT12) in dividing cells, which persisted with the same phase after the culture had entered the stationary growth phase. The relative velocities of the reactions catalyzed by the enzymes under both Km and saturating conditions was measured at different CTs, and their ratios were calculated.20 The values obtained for NADP phosphatase exhibited a complex pattern of rhythmicity, while those for NAD kinase displayed circadian variations strongly correlated with the rhythms in enzyme activity. The curves for showed troughs at CT00–04 both in dividing and nondividing cells and peaks at CT18–20 or at CT08–14 in cells sampled, respectively, from a dividing or a stationary culture. Such variations are indicative of changes in the kinetic properties of the enzyme, which may reflect modifications in its affinity either for effectors (such as Ca2+–calmodulin) or for its substrate, NAD+. This may be due to (i) the expression of different isoenzymes at different CTs; (ii) different posttranslational modifications of the enzyme; or (iii) concentrations of effectors varying in a circadian manner.
Adenylate Cyclase, Phosphodiesterase, Cyclic AMP and Cyclic GMP
A bimodal circadian rhythmicity (peaks at CT09–14, CT19–22) in total cyclic AMP (cAMP) content has been found in dividing and nondividing photoautotrophic cultures of wild-type E. gracilis Z under either entraining (LD:12,12) or free-running (LD:1/2,1/2,) regimes.72 These findings confirm that the bimodal rhythm of cAMP content is regulated by the CO and is not dependent on the CDC.
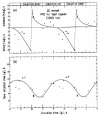
Similarly, cultures of the achlorophyllous ZC mutant, displaying free-running CRs of cell division in DD16 (cf. fig. 5), also exhibited bimodal circadian variations in their level of cAMP17 (fig. 10). Maximum cAMP levels occurred at the beginning of the light period (CT00–02, when cells are in G1) and at the onset of dark (CT12–14, corresponding to the onset of M). These variations appeared to be CDC-independent, since they persisted after cultures had reached the stationary phase (fig. 10B,C), although more recent results indicate that the oscillations in cAMP level may become attenuated in cell populations during late stationary-phase conditions.72 These oscillations in cAMP concentration could be phase-shifted by light signals17 in a manner predicted by the PRC previously derived for the phase-resetting of the division rhythm by light signals16 (fig. 5).
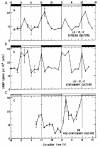
Key factors in the cAMP metabolic pathway are two enzymes responsible for its generation and degradation, namely, adenylate cyclase (AC) and phosphodiesterase (PDE). In LD:12,12 these enzymes were found to undergo bimodal, circadian variation of activity in both dividing and nondividing cultures of the ZC mutant of Euglena,21 although more recent results suggest that the amplitudes of the rhythms are damped in later-stationary-phase populations. Maximal AC activity occurred 2 h after the onset of the light interval (CT02) and at the beginning of darkness (CT12–14); these times corresponded to the acrophase profile for the rhythmic changes in cAMP content (cf. fig. 10) that have been reported previously.17 The activity of PDE also exhibited a daily oscillation, but with an inverse phase pattern. Both the AC and PDE activity rhythms persisted in DD (fig. 11).
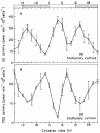
The activity of AC was activated significantly in vivo by forskolin,21 a highly specific activator of AC that can fully stimulate AC activity directly, independently of G proteins. Forskolin stimulation of AC did not result in uniform effects throughout the 24-h day but varied with CT. The degree of AC potentiation by an in vivo forskolin pulse appeared to be inversely dependent on the basal level of AC. Since forskolin appeared to stimulate AC activity at different CTs to the same maximal value regardless of its initial basal level, we conclude that the oscillatory activity of the enzyme derives from modulatory cellular effectors emanating from the CO rather than by changes in the amount of enzyme protein itself.
PDE is an important component of the signal system because it is responsible for the destruction of cAMP after a pulse of synthesis and, thereby, permits the cell to recover from a refractory state induced by cAMP. We found that the addition of 50 μM 3-isobutyl-1-methylxanthine (IBMX) to intact cells at CT20, corresponding to the peak of the rhythm in PDE activity (see fig. 11B), depressed PDE activity by about 51%, while at CT12 (the trough phase) IBMX had only a slight (8%) effect.21 These CT-dependent inhibitory effects of IBMX on PDE observed in Euglena extracts may be the resultant of its action on several PDE species, whose relative amounts may also vary with CT. These results indicate that the rhythms of both AC and PDE are key factors in generating the circadian oscillations of cAMP content.
Cyclic AMP and cyclic GMP (cGMP) are second messengers that may act in opposition as a dualistic “Yin Yang” type of system in biological regulation.101 To determine if cGMP plays a role in the mediation of circadian rhythmicity of the AC–cAMP–PDE system in Euglena, the levels of cAMP and cGMP were monitored in synchronized cell populations, and the effects of the cGMP analog 8-bromo-cGMP (8-Br-cGMP) and the cGMP inhibitor LY 83583 (6-anilinoquinone-5,8-quinone) on the activity of AC and PDE, as well as on the level of cAMP, were measured in vivo.22 A bimodal, 24-h rhythm of cGMP content was found in both dividing and nondividing cultures in either LD:12,12 or DD. The peaks and troughs of the cGMP rhythm occurred 2 h in advance of those of cAMP. The addition of 8-Br-cGMP at different CTs increased the cAMP level in vivo by two to five times, while LY 83583 reduced the amplitude of the cAMP rhythm so that it disappeared. The effects of 8-Br-cGMP on the activity of AC and PDE were CT-dependent, consistent with the changes in cAMP content. These findings suggest that cGMP may serve as an upstream effector that mediates the cAMP oscillation by regulation of the AC–cAMP–PDE system.
Coupling between Circadian Oscillator and Cell Cycle Clock
We have reviewed evidence for circadian clock control of the cell division cycle of Euglena (as well as many other microorganisms). This “gating” phenomenon is thought to reflect an interaction between a CO and the CDC. This coupling may be effected by cAMP, which has been implicated both in the regulation of the CDC and in circadian clock function, as a coupling link in the input and output pathways, if not as an element of the oscillator itself.18
The CDC of eukaryotes is a complex cascade of events that culminates in cell duplication. 102 These transitions from one regulatory state to another initiate the modification of substrates that determine the physical state of the cell and are themselves feedback-controlled. The basic mechanism, elucidated by genetic and biochemical analyses, has been conserved in organisms ranging from unicells such as yeasts, to Xenopus, clams, starfish eggs, cultured human cells and higher plants (reviewed in ref. 103).
Two crucial transition control points exist at the G1/S and G2/M boundaries of the CDC. The M phase is characterized by the activation of a kinase [MPF, or Maturation (M phase)-Promoting Factor, so named for its ability to induce cell division when injected into Xenopus oocytes], which consists of two component subunits. One is the p34cdc2 protein (the product of the S. pombe gene cdc2, or its homologue, CDC28, in S. cerevisiae), which is able to phosphorylate casein and histone H1 in vitro and has maximal kinase activity at mitosis. The other is cyclin, whose cellular concentration fluctuates during the CDC. It accumulates gradually during interphase, forms a complex with p34cdc2, and activates the protein kinase; it is then degraded during mitosis, turning off kinase and MPF activity. The cyclin–cdc2/CDC28 (MPF) system thus behaves like an oscillator, or “clock,” which is reset to its interphase state during mitosis and which appears to constitute a universal cell-division-cycle “engine.”103 We now know that there are entire families of cyclins with which p34cdc2 periodically associates to create successive waves of protein kinase activity that regulate the progression of the CDC and that distinct classes of cdc2-related genes are differentially expressed during the CDC in plants and animals.
Role of Cyclic AMP
Cyclic AMP, which is known to play a pivotal role in cellular regulation,104 may effect a coupling between the CO and the p34cdc2–cyclin pathway, participating (perhaps in concert with other players) in the gating of CDC events to specific phases of the circadian cycle.
Indeed, cAMP seems to have the capacity to control certain rate-limiting steps in CDC progression, stimulating the proliferation of some cells, but having the opposite effect, or no effect at all, on others.105,106 Transient increases of cAMP are correlated with cell cycle transitions at both the G1/S and G2/M boundaries in many cell types. Genetic experiments in Saccharomyces,107 and pharmacological studies in mammalian cells,104 have shown that a transient rise and the ensuing fall in the cAMP level are necessary for the initiation of DNA synthesis and the tyrosine-dephosphorylation-induced activation of MPF,109 and that a second cAMP surge is correlated with the completion of, and exit from mitosis.110 Similar changes of cAMP concentration at different phases of the CDC have been shown to occur in Euglena.111 These signals coincide with the action(s), respectively, of p34cdc2–p60 (G1 cyclin) protein kinase complexes and of p34cdc2–cyclin B (mitosis cyclin) protein kinase complexes.
Thus, fluctuations in cAMP concentration represent an important link in signal transduction during CDC progression in animal and yeast cells. In eukaryotes, transcriptional regulation upon stimulation of the adenylyl cyclase signaling pathway is mediated by a family of cAMP-responsive nuclear factors. These factors contain the basic domain/leucine zipper motifs and bind as dimers to cAMP-response elements (CRE). The function of CRE-binding proteins (CREB) is modulated by phosphorylation by the cAMP-dependent protein kinase.
Circadian Rhythmicity in Cyclic AMP Levels
We have discussed the CDC-independent, bimodal CRs of cAMP levels observed in cultures of both wild-type Euglena50,51 and of the achlorophyllous ZC mutant52 (cf. fig. 10) displaying free-running CRs of cell division. Maximum cAMP levels occurred at the beginning of the light period (CT00–02, when cells are in G1) and at the onset of dark (CT12–14, corresponding to the onset of M). Similar fluctuations of cAMP that are tightly connected to cell cycle progression in synchronized suspension cultures of tobacco BY-2 have recently been reported. 112 Cyclic AMP peaks were observed during the G1 and S phases. Application of indomethacin, a drug that inhibits adenylyl cyclase in animal cells, at early S phase resulted in the loss of the S-phase cAMP peak and inhibited mitotic division by blocking cells at the G2/M border.
Collectively, these results suggest that cAMP levels are controlled by the same endogenous clock as that regulating the CDC and that the periodic, bimodal cAMP signal (initiated by the CO, or in conjunction with it) may participate in the gating of CDC events to specific phases of the circadian cycle.
Perturbation of the Cell Division Rhythm by Cyclic AMP
If periodic cAMP signals play a role in the generation of cell division rhythmicity in Euglena, then conditions that disturb or override the the control of the cAMP level by the CO should also cause derangements in downstream events.18 Perturbations of the cAMP oscillation (see fig. 10) by exogenous cAMP resulted in the temporary uncoupling of the CDC from the circadian timer, causing either a shortening or a lengthening of the CDC, depending on the phase of the CDC when the drug was applied (fig. 12A). Delays of the next synchronous division step (up to 9 h) were obtained when cAMP was given between CT03 and CT09, and advances were observed when cAMP was given between CT16 and CT22. Maximum effects were obtained at CT06–08 and CT18–20, corresponding to times when endogenous cAMP levels were minimal (see fig. 10). Dose-response curves were also derived at CTs when maximum +ΔΦs or -ΔΦs of the CDC had been obtained. As little as 1 nM cAMP was enough to perturb CDC transit. The CO, in contrast to the CDC cyclin clock, and unlike excitable tissue in which cAMP does phase-shift the output rhythm,113,114 was not reset by addition of cAMP: the division rhythm returned to its original phase (fig. 12B).
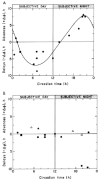
The ΔΦs of the division steps observed in the growth curves following cAMP injection reflected real changes in the rate of CDC progression. Measurement of cell DNA content by flow cytometry18 indicated that cAMP injected at CT06–08 delayed progress through S phase, and perhaps through M (fig. 13A). If added at CT18–20, cAMP accelerated the G2/M transition (fig. 13B). These effects on the cell division rhythm were only transitory: After 48 h division occurred at the same phase as in unperturbed controls (see fig. 12B).

That the endogenous circadian variations of cAMP level were of sufficient amplitude to have similar effects on CDC progression was demonstrated by testing the effect of drugs that reduce the amplitude of the cAMP oscillation, or that keep cAMP at a level such that all cAMP receptors should be permanently saturated: We would expect such drugs to prevent the expression of division rhythmicity. Indeed, when the cAMP analog forskolin, an activator of AC that maintained cAMP at an abnormally high level,21 was added to a culture of the ZC mutant free-running in DD, a rapid desynchronization of the cell population occurred and division rhythmicity eventually was lost.18
These findings indicate that although cAMP signals do not reset the CO in Euglena (thus, cAMP is unlikely to represent a “gear” of the clock), they do regulate CDC progression, acting downstream from the oscillator—perhaps in a parallel pathway, as Zatz115 has suggested for chick pineal cells.
Downstream Pathway: Cyclic AMP-Dependent Kinases
What are the coupling links between the oscillatory cAMP system and the CDC? Whatever be the cell cycle regulatory pathways that are affected, one must explain how identical signals can perturb different pathways depending on the CT at which they are applied, so that cell cycle delays are obtained during the subjective days, and advances during the subjective nights. One possibility is that cAMP acts through different “receptors” that selectively modulate one or the other of the two regulatory pathways.
We have demonstrated that the ZC mutant of Euglena contains two types of cAMP-dependent kinase (cPKA and cPKB), which have different affinities for cAMP and for several cAMP analogs.116 Cell extracts were found to contain two cAMP-binding proteins, which bound cAMP with a high affinity (Kd values of 10 nM and 30 nM) and which could be separated by DEAE-cellulose chromatography. Protein kinase activity was assayed using Kemptide as a substrate (specifically phosphorylated by cAMP-dependent kinases from mammalian cells). Stimulation of kinase activity by cAMP was observed after partial purification by DEAE-cellulose chromatography. Two peaks of activity were resolved, corresponding to enzymes with different cAMP-analog specificities. Thus, cAMP signalling in plant cells may proceed by phosphorylation of target proteins by cAMP-dependent kinases in a manner similar to that of animal cells.
The differential activation of the two kinases identified in Euglena extracts by cAMP analogs afforded a tool for the study of their respective roles in the control of CDC progression; we determined the mimimum doses of cAMP or of cAMP analogs that caused ΔΦs of the division rhythm during the subjective day, or during the subjective night.18 Different results were obtained at CT06–08, and at CT18–20, suggesting that the effect of cAMP at the different CTs was mediated by two different cAMP kinases. Thus, 8-benzylamino-cAMP (8-BZA-cAMP), which selectively activates cPKA, induced -ΔΦs when added at CT06–08, but had no effect at CT18–20. Reciprocally, 8-(4-chlorophenylthio)-cAMP (8-CPT-cAMP), which specifically activates cPKB, induced +ΔΦs and a loss of division rhythmicity when added at CT18–20 but did not perturb the division rhythm when added at CT06–08.
There also was a correlation between the doses of cAMP, 8-BZA-cAMP, 8-CPT cAMP, and N6 -monobutyryl-cAMP (6-MBT-cAMP) that caused perturbations of the CDC at CT06–08 and the Ka values of cPKA for these analogs, a finding that suggests that cPKA mediates the delaying effects of cAMP at these CTs. Similarly, there was a correlation between the doses of the same nucleotides that caused perturbations of the CDC at CT18–20 and the Ka of cPKB for these analogs, a result indicating that cPKB mediates the accelerating effects of cAMP at CT18–20. A simple explanation for these findings would be that these kinases are expressed at different stages of the CDC, as has been described for type I and II cAMP-dependent kinases in mammalian cells.
Model for Circadian Control of the Cell Division Cycle
These results were incorporated into a model (fig. 14) for the coupling of the CDC to the CO.18 We have proposed that the cAMP surge at CT00–02 delays DNA synthesis and then holds the cells at a restriction point in G2 to prevent cell division during the subjective day. The cells are released from this blockage after cAMP levels subside, which Grieco et al119 have shown is associated with the tyrosine-dephosphorylation-induced activation of MPF, and the G2/M transition, or mitosis itself, is accelerated by the second cAMP peak, at CT12–14, so that mitosis is phased to the subjective night. The delaying effects of cAMP on CDC progression during the subjective day would be mediated by the activation of cPKA, and the stimulation of mitosis during the subjective night by the activation of cPKB. Activation of either of these kinases would cause the phosphorylation of a different set of targets and perturb different cell-cycle control pathways. cPKA and cPKB may be expressed at different phases of the CDC. Alternatively, the level of these enzymes might exhibit circadian variations, with cPKA's being expressed during the subjective day and cPKB during the subjective night. Another possibility is that the level of one of their downstream targets oscillates, so that only cPKA activation has an effect on CDC progression during the subjective day, and cPKB during the subjective night.
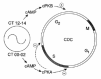
Interface with the Cyclin Oscillator
How does the CO, acting via (or in conjunction with) the AC–cAMP–PDE system and the cAMP-dependent kinases (cPKA and cPKB) that we have identified, ultimately couple to the MPF clock regulating the CDC? Recently, we have run western blot analyses of cyclin B (the regulatory subunit of MPF) using monoclonal antibody to human clb1 protein and have found that the abundance of the homolog in protein extracts from dividing ZC cultures varied quantitatively during both the circadian cycle and the CDC,73,74 with a peak at CT00 and a trough at CT12. These circadian fluctuations in cyclin B in dividing cells ceased, however, during the nondividing phase in stationary-phase cultures.
Preliminary western blot analyses of protein extracts from dividing and stationary-phase cultures of the ZC mutant in DD73,74 confirm the presence of a homolog of p34cdc2, the catalytic subunit of MPF, reported earlier in wild type Euglena and in other algae117,118 and suggest that its abundance throughout the CDC is invariant, decreasing to a much reduced level when cells cease dividing. Further, a relative shift in electrophoretic mobility of the monomer was observed to occur in a cycle-dependent manner: A more slowly moving band disappeared at CT09 and reappeared at CT18. Such changes in mobility have been attributed to phosphorylation and dephosphorylation. Remarkably, this rhythm of phosphorylation of cdc2 persisted with a circadian period (τ = 26h), even in stationary-phase cultures. We determined the nature of these changes by using potato acid phosphatase to completely dephosphorylate cdc2, which then ran as a single band with no anti-phosphotyrosine immunoreactivity.73,74 These initial findings suggest that the activity of this polypeptide depends on posttranslational mechanisms.
More recent results80 using untreated extracts have revealed, in addition to dephosphorylated protein, two phosphorylated forms of cdc2. The more slowly moving (more phosphorylated) form predominated during the subjective day and was the only form recognized by anti-phosphotyrosine. The faster moving, less phosphorylated form appeared during the subjective night concomitant with a decline in the more slowly moving form. The faster moving, phosphorylated form lacked tyrosine phosphorylation and, in fact, may be threonine-phosphorylated, as is the case in other species. By the end of the subjective night, however, this form disappeared, while the amount of the more slowly moving form, now phosphorylated at both residues, became elevated before subjective dawn. We conclude that reversible tyrosine phosphorylation of cdc2 is coupled to the CO.
Finally, we note that Bjarnason and coworkers119,120 have examined the circadian expression of clock genes in human oral mucosa and skin and their association with specific CDC phases. The relative RNA expression of hClock, hTim, hPer1, hCry1 and hBmal1 was determined in biopsies taken throughout one timespan of 24 h and was found to exhibit a circadian profile consistent with that found in the SCN and peripheral tissue of rodents. hPer1, hCry1 and hBmal1 were rhythmic, peaking in the early morning, late afternoon, and at night, respectively, whereas hClock and hTim were arrhythmic. In concurent oral mucosa biopsies, thymidylate synthase activity (a marker for DNA synthesis), had a circadian variation with an early afternoon peak, coinciding with the timing of S phase. The major peak in hPer1 expression occurred at the same time of day as the peak in the G1 phase in oral mucosa, suggesting a link between the CO and the mammalian CDC. Similarly, the CDC proteins, p53, cyclin E, cyclin A and cyclin B1 were found to vary rhythmically, with their respective peaks' occurring in late G1, G1/S, G2 and M. Likewise, in an analysis of some 68 CDC-related genes in regenerating liver of mice, using DNA microarray and Northern blot techniques, Matsuo et al121 have shown that the circadian clock directly controls the expression of wee1, which in turn modulates the expression of active cyclin B1–Cdc2 kinase and the entry of cells into mitosis. These findings have important clinical implications for the timing of both chemotherapy and radiotherapy, as illustrated by a recent study122 reporting that proapoptotic drugs such as docetaxel displayed least toxicity and highest antitumor efficacy following dosing during the circadian rest phase in mice, a finding that suggests that the CDC and apoptotic processes (e.g., expression of the proapoptotic BCL 2-associated X protein) could be regulated by the circadian clock.
Conclusion
From the foregoing review, it is clear that the Euglena system has continued to provide an excellent case for temporal differentiation. In addition to the numerous examples cited in major reviews some 20 years ago,2,24 many new instances of diverse behavorial, physiological and biochemical activities that are partitioned along the 24-hour time axis have been documented by several laboratories. These include the demonstration of the photoperiodic control of the CDC, the presence of melatonin, and the circadian variation in light-harvesting chlorophyll proteins, NAD+ kinase and NADP phosphatase (with implications for the mitochondrial calcium cycle), and the entire adenylate cyclase—cAMP—phosphodiesterase pathway (with a tie-in to the cyclin–p34cdc2/CDC28 universal cell-division-cycle “engine”). Although Euglena lacks sexuality and, therefore, is not amenable to the genetic dissection afforded by such model systems as Neurospora, Drosophila, and Arabidopsis, it has the advantage of a versatility in nutritional modes, the ease with which it can be cultured and its CDC can be synchronized, and the facility with which mutants can be obtained for analysis of the chloroplast compartment and its contribution to various physiological and biochemical phenomena. These considerations have enabled and abetted the experimental studies of circadian clocks reported herein and should continue to do so.
References
- 1.
- Edmunds JrLN. Temporal differentiation in Euglena: Circadian phenomena in nondividing populations and in synchronously dividing cellsIn: Lefort-Tran M, Valencia R, eds.Les Cycles Cellulaires et leur Blocage chez Plusieurs Protistes. Colloques Int CNRS, No 240Paris: Centre National de la Recherche Scientifique,197553–67.
- 2.
- Edmunds JrLN. Circadian and infrdian rhythmsIn: Buetow DE, ed.The Biology of Euglena. Vol IIINew York: Academic Press,198253–142.
- 3.
- Edmunds JrLN, Halberg F. Circadian time structure of Euglena: A model system amenable to quantificationIn: Kaiser HE, ed.Neoplasms—Comparative Pathology of Growth in Animals, Plants and ManBaltimore: Williams & Wilkins Co,1981105–134.
- 4.
- Wille JrJJ. Biological rhythms in protozoaIn: Levandowsky M, Hutner SH, eds.Biochemistry and Physiology of Protozoa. 2nd ed., vol.2New York: Academic Press,1979267–149.
- 5.
- Lloyd D. Circadian and ultradian clock-controlled rhythms in unicellular microorganisms. Adv Microb Physiol. 1998;39:291–338. [PubMed: 9328650]
- 6.
- Cook JR, James TW. Light-induced division synchrony in Euglena gracilis var. bacillaris. Exp Cell Res. 1960;21:583–589. [PubMed: 13695255]
- 7.
- Edmunds JrLN. Studies on synchronously dividing cultures of Euglena gracilis Klebs (strain Z). I. Attainment and characterization of rhythmic cell division. J Cell Comp Physiol. 1965;66:147–158. [PubMed: 5862681]
- 8.
- Terry OW, Edmunds JrLN. Phasing of cell division by temperature cycles in Euglena cultured autotrophically under continuous illumination. Planta. 1965;93:106–127. [PubMed: 24496707]
- 9.
- The Biology of Euglena. Vol IIn: Buetow DE, ed. New York: Academic Press,1968 .
- 10.
- The Biology of Euglena. Vol IIIn: Buetow DE, ed.New York: Academic Press,1968 .
- 11.
- The Biology of Euglena. Vol IIIIn: Buetow DE, ed. New York: Academic,1982 .
- 12.
- Kirschstein M. Das rhythmische Verhalten einer farblosen Mutante von Euglena gracilis. Planta. 1969;85:126–134. [PubMed: 24515585]
- 13.
- Jarrett RM, Edmunds JrLN. Persisting circadian rhythm of cell division in a photosynthetic mutant of Euglena. Science. 1979;167:1730–1733. [PubMed: 5416537]
- 14.
- Mitchell JLA. Photoinduced division synchrony in permanently bleached Euglena gracilis. Planta. 1971;100:244–257. [PubMed: 24488197]
- 15.
- Edmunds JrLN, Jay ME, Kohlmann A. et al. The coupling effects of some thiol and other sulfur-containing compounds on the circadian rhythm of cell division in photosynthetic mutants of Euglena. Arch Microbiol. 1976;108:1–8. [PubMed: 818971]
- 16.
- Carré I, Oster AS, Laval-Martin DL. et al. Entrainment and phase shifting of the circadian rhythm of cell division by light in cultures of the achlorophyllous ZC mutant of Euglena gracilis. Curr Microbiol. 1989;19:223–229.
- 17.
- Carré I, Laval-Martin DL, Edmunds JrLN. Circadian changes in cyclic AMP levels in synchronously dividing and stationary-phase cultures of the achlorophyllous ZC mutant of Euglena gracilis. J Cell Sci. 1989;94:267–272. [PubMed: 1661740]
- 18.
- Carré IA, Edmunds JrLN. Oscillator control of cell division in Euglena: Cyclic AMP oscillations mediate the phasing of the cell division cycle by the circadian clock. J Cell Sci. 1993;104:1163–1173. [PubMed: 8391014]
- 19.
- Laval-Martin DL, Carré I, Barbera SJ. et al. Rhythmic changes in the activities of NAD kinase and NADP phosphatase in the achlorophyllous ZC mutant of Euglena. Arch Biochem Biophys. 1990;276:433–441. [PubMed: 2154948]
- 20.
- Laval-Martin DL, Carré I, Barbera SJ. et al. Circadian variations in the affinities of NAD kinase and NADP phosphatase for their substrates, NAD+ and NADP+, in dividing and nondividing cells of the achlorophyllous ZC mutant of Euglena gracilis (strain Z). Chronobiol Int. 1990;7:99–105. [PubMed: 2173644]
- 21.
- Tong J, Carré IA, Edmunds JrLN. Circadian rhythmicity in the activities of adenylate cyclase and phosphodiesterase in synchronously dividing and stationary-phase cultures of the achlorophyllous ZC mutant of Euglena. J Cell Sci. 1991;100:365–369. [PubMed: 1661740]
- 22.
- Tong J, Edmunds JrLN. Role of cyclic GMP in the mediation of circadian rhythmicity of the adenylate cyclase–cyclic AMP–phosphodiesterase system in Euglena. Biochem Pharmacol. 1993;45:2087–2091. [PubMed: 8390260]
- 23.
- Wolff D, Künne A. Light-regulated, circadian respiration activity of Euglena gracilis mutants that lack chloroplasts. J Plant Physiol. 2000;156:52–59.
- 24.
- Edmunds JrLN. Physiology of circadian rhythms in microorganismsIn: Rose AH, Tempest DW, eds.Advances in Microbial PhysiologyLondon: Academic Press,19842561–148. [PubMed: 6398623]
- 25.
- Edmunds JrLN. Replication of DNA and cell division in synchronously dividing cultures of Euglena gracilis. Science. 1964;145:266–268. [PubMed: 14171566]
- 26.
- Edmunds JrLN. Clocked cell cycle clocks: Implications toward chronopharmacology and agingIn: Samis Jr HV, Capobianco S, eds.Aging and Biological RhythmsNew York: Plenum,1978125–184. [PubMed: 742525]
- 27.
- Edmunds JrLN, Funch RR. Circadian rhythm of cell division in Euglena: Effects of a random illumination regimen. Science. 1969;165:500–503. [PubMed: 5793244]
- 28.
- Edmunds JrLN, Funch RR. Effects of ‘skeleton’ photoperiods and high frequency light–dark cycles on the rhythm of cell division in synchronized cultures of Euglena. Planta. 1969;87:134–163. [PubMed: 24504723]
- 29.
- Edmunds JrLN, Laval-Martin DL. Cell division cycles and circadian oscillatorsIn: Edmunds Jr LN, ed.Cell Cycle ClocksNew York: Marcel Dekker,1984295–324.
- 30.
- Hagiwara S-Y, Takahashi M, Yamagishi A. et al. Novel findings regarding photoinduced commitments of G1-, S- and G2-phase cells to cell-cycle transitions in darknes and dark-induced G1-, Sand G2-phase arrests in Euglena. Photochem Photobiol. 2001;74:726–733. [PubMed: 11723802]
- 31.
- Hagiwara S-Y, Bolige A, Zhang Y. et al. Circadian gating of photoinduction of commitment to cell-cycle transitions in relation to photoperiodic control of cell reproduction in Euglena. Photochem Photobiol. 2002;76:105–115. [PubMed: 12126300]
- 32.
- Edmunds JrLN. Persisting circadian rhythm of cell division in Euglena: Some theoretical considerations and the problem of intercellular communicationIn: Menaker M, ed.BiochronometryWashington, DC: National Academy of Sciences,1971594–611.
- 33.
- Edmunds JrLN, Chuang L, Jarrett RM. et al. Long-term persistence of free-running rhythms of cell division in Euglena and the implication of autosynchrony. J Interdisc Cycle Res. 1971;2:121–132.
- 34.
- Brinkmann K. Temperatureinflüsse auf die circadiane Rhythmik von Euglena gracilis bei Mixotrophie und Autotrophie. Planta. 1966;70:344–389. [PubMed: 24558006]
- 35.
- Schnabel G. Der Einflüss von Licht auf die circadiane Rhythmik von Euglena gracilis bei Autotrophie und Mixotrophie. Planta. 1968;81:49–63. [PubMed: 24519596]
- 36.
- Brinkmann K. Metabolic control of temperature compensation in the circadian rhythm of Euglena gracilisIn: Menaker M, ed.BiochronometryWashington, DC: National Academy of Sciences,1971567–593.
- 37.
- Kreuels T, Brinkmann K. The versatility of D2O effects on biological and chemical oscillations. Chronobiologia. 1979;6:121–122.
- 38.
- Kreuels T, Jörres R, Martin W. et al. System analysis of the circadian rhythm of Euglena gracilis, II: Masking effects and mutual interactions of light and temperature responses. Z Naturforsch. 1984;39c:801–811.
- 39.
- Martin W, Jörres R, Kreuels T. et al. Systemanalyse der circadianen Rhythmik von Euglena gracilis: Linearitäten und Nichtlinearitäten in der Reaktion auf Temperatursignale. Ber Deutsch Bot Ges. 1985;98:173–186.
- 40.
- Brinkmann K. Circadian rhythm in the kinetics of acid denaturation of cell membranes of Euglena gracilis. Planta. 1976;129:221–227. [PubMed: 24430960]
- 41.
- Walther WG, Edmunds JrLN. Studies on the control of the rhythm of photosynthetic capacity in synchronized cultures of Euglena gracilis (Z). Plant Physiol. 1973;51:250–258. [PMC free article: PMC366245] [PubMed: 16658310]
- 42.
- Laval-Martin DL, Shuch DJ, Edmunds JrLN. Cell cycle-related and endogenously controlled circadian photosynthetic rhythms in Euglena. Plant Physiol. 1979;63:495–502. [PMC free article: PMC542858] [PubMed: 16660755]
- 43.
- Edmunds JrLN, Laval-Martin DL. ‘Free-running’ circadian rhythms of photosynthesis elicited by short-period cycles of light and darkness in synchronously dividing and nondividing EuglenaIn: Akoyunoglou G, ed.Photosynthesis. Vol II. (Proc 5th Int Cong on Photosynthesis, Halkidiki, Greece, 1980)Philadelphia: Balaban International Science Services,1981313–322.
- 44.
- Lonergan TA, Sargent ML. Regulation of the photosynthesis rhythm in Euglena gracilis. I. Carbonic anhydrase and glyceraldehyde-3-phosphate dehydrogenase do not regulate the photosynthesis rhythm. Plant Physiol. 1978;61:150–153. [PMC free article: PMC1091822] [PubMed: 16660250]
- 45.
- Lonergan TA, Sargent ML. Regulation of the photosynthesis rhythm in Euglena gracilis. II. Involvement of electron flow through both photosystems. Plant Physiol. 1979;64:99–l03. [PMC free article: PMC543032] [PubMed: 16660924]
- 46.
- Lonergan TA. Regulation of cell shape in Euglena gracilis. I. Involvement of the biological clock, respiration, photosynthesis, and cytoskeleton. Plant Physiol. 1983;71:719–730. [PMC free article: PMC1066111] [PubMed: 16662896]
- 47.
- Lonergan TA. A possible second role for calmodulin in biological clock-controlled processes of Euglena. Plant Physiol. 1986;82:226–229. [PMC free article: PMC1056094] [PubMed: 16664997]
- 48.
- Lonergan TA. The photosynthesis and cell shape rhythms can be naturally uncoupled from the biological clock in Euglena gracilis. J Exp Bot. 1986;37:1334–1340.
- 49.
- Pohl R. Tagesrhythmus in phototakischen verhalten der Euglena gracilis. Z Naturforsch. 1948;3b:367–374.
- 50.
- Bruce VG, Pittendrigh CS. Temperature independence in a unicellular “clock” Proc Natl Acad Sci USA. 1956;42:676–681. [PMC free article: PMC534275] [PubMed: 16589930]
- 51.
- Bruce VG, Pittendrigh CS. Resetting the Euglena clock with a single light stimulus. Am Nat. 1958;92:294–306.
- 52.
- Feldman JF. Lengthening the period of a biological clock in Euglena by cycloheximide, an inhibitor of protein synthesis. Proc Natl Acad Sci USA. 1967;57:1080–1087. [PMC free article: PMC224658] [PubMed: 5231347]
- 53.
- Feldman JF, Bruce VG. Circadian rhythm changes in autotrophic Euglena induced by organic carbon sources. J Protozool. 1972;19:370–372. [PubMed: 4624304]
- 54.
- Balzer I, Neuhaus-Steinmetz U, Quentin E. et al. Concomitance of circadian and circa-4-hour ultradian rhythms in Euglena gracilis. J Interdisc Cycle Res. 1989;20:15–24.
- 55.
- Terry OW, Edmunds JrLN. Rhythmic settling induced by temperature cycles in Euglena cultured autotrophically under continuous illumination. Planta. 1970;93:128–142. [PubMed: 24496707]
- 56.
- Kiefner G, Schliessmann F, Engelmann W. Methods for recording circadian rhythms in Euglena. Int J Chronobiol. 1974;2:189–195.
- 57.
- Lonergan TA. Regulation of cell shape in Euglena gracilis. II. The effects of altered extra- and intracellular Ca2+ concentrations and the effect of calmodulin antagonists. J Cell Sci. 1984;71:37–50. [PubMed: 6440896]
- 58.
- Lonergan TA. Regulation of cell shape in Euglena gracilis. IV. Localization of actin, myosin and calmodulin. Cell Sci. 1985;77:197–208. [PubMed: 3936854]
- 59.
- Lachney CL, Lonergan TA. Regulation of cell shape in Euglena gracilis. III. Involvement of stable microtubules. J Cell Sci. 1985;74:219–237. [PubMed: 3928636]
- 60.
- Brinkmann K. The influence of alcohols on the circadian rhythm and metabolism of Euglena gracilis. J Interdisc Cycle Res. 1976;7:149–170.
- 61.
- Cook JR. Euglena gracilis in synchronous division. II. Biosynthetic rates over the light cycle. Biol Bull. 1961;121:277–289.
- 62.
- Edmunds JrLN. Studies on synchronously dividing cultures of Euglena gracilis Klebs (strain Z). II. Patterns of biosynthesis during the cell cycle. J Cell Comp Physiol. 1965;66:159–182. [PubMed: 5862682]
- 63.
- Feldman JF. Circadian rhythmicity in amino acid incorporation in Euglena gracilis. Science. 1968;160:1454–1456. [PubMed: 5654056]
- 64.
- Künne A, de GrootEJ. Protein synthesis in Euglena gracilis is light- and temperature-dependent, oscillating in a circadian, temperature-compensated manner. Bot Acta. 1996;109:57–63.
- 65.
- Sulzman FM, Edmunds JrLN. Persisting circadian oscillations in enzyme activity in nondividing cultures of Euglena. Biochem Biophys Res Commun. 1972;47:1338–1344. [PubMed: 4625140]
- 66.
- Sulzman FM, Edmunds JrLN. Characterization of circadian oscillations in alanine dehydrogenase activity in nondividing populations of Euglena gracilis (Z). Biochim Biophys Acta. 1973;320:594–609. [PubMed: 4201692]
- 67.
- Edmunds JrLN, Sulzman FM, Walther WG. Circadian oscillations in enzyme activity in Euglena and their relation to the circadian rhythm of cell divisionIn: Scheving LE, Halberg F, Pauly JE, eds.ChronobiologyTokyo: Igaku Shoin,197461–66.
- 68.
- Goto K, Laval-Martin D, Edmunds JrLN. Biochemical modeling of an autonomously oscillatory circadian clock in Euglena. Science. 1985;228:1284–1288. [PubMed: 2988128]
- 69.
- Pfleging P, Morawietz G, Hardeland R. Differences in oscillation frequencies between two key enzymes of aromatic amino acid catabolism in Euglena: Circadian rhythmicity of trytophan 2,3-dioxygenase vs. predominance of ultradian periods in tyrosine aminotransferase. Biol Rhythm Res. 1994;25:59–66.
- 70.
- Quentin E, Hardeland R. Circadian rhythmicity of protein synthesis and translational control in Euglena gracilis. J Interdisc Cycle Res. 1986;17:197–205.
- 71.
- Quentin E, Hardeland R. Stimulation of 80 S protein synthesis by heat-stable cytosolic extracts in Euglena gracilis. Comp Biochem Physiol. 1986;85B:89–92.
- 72.
- Mohabir G, Edmunds JrLN. Circadian clock regulation of the bimodal rhythm of cyclic AMP in wild-type Euglena. Cell Signal. 1999;11:143–147. [PubMed: 10048792]
- 73.
- Edmunds JrLN, Mohabir G. Regulation of the cell division cycle in Euglena gracilis: Role of reversible tyrosine phosphorylation in the timing of mitotic kinase activity. Physiologist. 1995;38(3):A–19.
- 74.
- Mohabir G, Edmunds JrLN. Reversible phosphorylation of mitotic kinase (CDC2) is under circadian control in the unicell Euglena gracilis. Biol Rhythm Res. 1995;26:420–42l.
- 75.
- Edmunds JrLN. Cross-talk between clocks: Regulation of cell division cycles by circadian oscillatorsIn: Vanden Driessche T, Guisset J-L, Petiau-de Vries G, eds.Membranes and Circadian RhythmsBerlin, Heidelberg: Springer-Verlag,199695–124.
- 76.
- Künne A, Pistorius E, de GrootE. Characterization of polypeptides in Euglena gracilis which are synthesized in a circadian manner. Eur J Cell Biol. 1997;73:175–181. [PubMed: 9208231]
- 77.
- Künne A, Pistorius E, Kloppstech K. et al. Circadian synthesis of light-harvesting-chlorophyll-proteins in Euglena gracilis is under translational control. Z Naturforsch. 1998;53c:1917–1026. [PubMed: 9933967]
- 78.
- Balzer I, Fuhrberg B. Presence of melatonin in the unicell Euglena gracilis. (Abstracts, 5th Mtg Society for Research on Biological Rhythms, 8-12 May) Jacksonville, Florida: Amelia Island Plantation 199682(#111)
- 79.
- Balzer I, Höcker B, Kapp H. et al. Occurrence and comparative physiology of melatonin in evolutionary diverse organismsIn: Vanden Driessche T, Guisset J-L, Petiau-de Vries G, eds.The Redox State and Circadian RhythmsDordrecht: Kluwer Academic Publishers,200095–119.
- 80.
- Edmunds JrLN, Mohabir G. Distinct phosphorylated forms of cdc2, alternating with a circadian period in free-running Euglena, persist in cultures arrested by carbon starvation. (Abstracts, 5th Mtg Society for Research on Biological Rhythms, 8-12 May) Jacksonville, Florida: Amelia Island Plantation 199644(No35)
- 81.
- Ledoigt G, Calvayrac R. Phénomènes périodiques, métaboliques et structuraux chez un protiste, Euglena gracilis. J Protozool. 1979;26:632–643. [PubMed: 94608]
- 82.
- Edmunds JrLN. Studies on synchronously dividing cultures of Euglena gracilis Klebs (strain Z). III. Circadian components of cell division. J Cell Physiol. 1966;67:35–44. [PubMed: 5937015]
- 83.
- Edmunds JrLN, Tay DE, Laval-Martin DL. Cell division cycles and circadian clocks: Phase-response curves for light perturbations in synchronous cultures of Euglena. Plant Physiol. 1982;70:297–302. [PMC free article: PMC1067129] [PubMed: 16662464]
- 84.
- Malinowski JR, Laval-Martin DL, Edmunds JrLN. Circadian oscillators, cell cycles, and the singularity: Light perturbations of the free-running rhythm of cell division in Euglena. J Comp Physiol. 1985;B155:257–267. [PubMed: 3837018]
- 85.
- Anderson RW, Laval-Martin DL, Edmunds JrLN. Temperature compensation of the circadian rhythm of cell division in Euglena. Exp Cell Res. 1985;157:144–158. [PubMed: 3918876]
- 86.
- Winfree AT. Integrated view of resetting a circadian clock. J Theor Biol. 1970;28:327–374. [PubMed: 5487648]
- 87.
- Winfree AT. The Geometry of Biological TimeNew York: Springer-Verlag,1980 .
- 88.
- Edmunds JrLN. Cellular and Molecular Bases of Biological ClocksNew York: Springer-Verlag,1988 .
- 89.
- Sweeney BM, Hastings JW. Effects of temperature upon diurnal rhythms. Cold Spring Harb Symp Quant Biol. 1960;25:87–104. [PubMed: 13774256]
- 90.
- Spudich J, Sager R. Regulation of Chlamydomonas cell cycle by light and dark. J Cell Biol. 1980;85:136–145. [PMC free article: PMC2110596] [PubMed: 6767730]
- 91.
- Donnan L, John PCL. Cell cycle control by timer and sizer in Chlamydomonas. Nature (London). 1983;304:630–633. [PubMed: 6877383]
- 92.
- Yee M-C, Bartholomew JC. Effects of 3-(3,4-dichlorophenyl)-1,1-dimethylurea on the cell cycle in Euglena gracilis. Plant Physiol. 1989;91:1025–1029. [PMC free article: PMC1062112] [PubMed: 16667105]
- 93.
- Zachleder V, van den Ende H. Cell cycle events in the green alga Chlamydomonas eugametos and their control by environmental factors. J Cell Sci. 1992;102:469–474.
- 94.
- Krupinska K, Humbeck K. Light-induced synchronous cultures, an excellent tool to study the cell cycle of unicellular green algae. J Photochem Photobiol B. 1994;26:217–231.
- 95.
- Bostock CJ. DNA synthesis in the fission yeast Schizosaccharomyces pombe. Exp Cell Res. 60:16–26. [PubMed: 5439575]
- 96.
- Costello G, Rodgers L, Beach D. Fission yeast enters the stationary phase G0 state from either mitotic G1 or G2. Curr Genet. 1986;11:119–125.
- 97.
- Wei W, Nurse P, Broek D. Yeast cells can enter a quiescent state through G1, S, G2, or M phase of the cell cycle. Cancer Res. 1993;53:1867–1870. [PubMed: 8467507]
- 98.
- Tyson JJ, Novak B. Regulation of the eukaryotic cell cycle: Molecular antagonism, hysteresis, and irreversible transitions. J Theor Biol. 2001;210:249–263. [PubMed: 11371178]
- 99.
- Edmunds JrLN, Adams KJ. Clocked cell cycle clocks. Science. 1981;211:1002–1013. [PubMed: 7008196]
- 100.
- Goto K. Causal relationships among metabolic circadian rhythms in Lemna. Z Naturforsch. 1984;39c:73–84.
- 101.
- Goldberg ND, Haddox MK, Dunham E. et al. The Yin Yang hypothesis of biological control: Opposing influences of cyclic GMP and cyclic AMP in the regulation of cell proliferation and other biological processesIn: Clarkson B, Baserga R, eds.Control of Proliferation in Animal CellsCold Spring Harbor: Cold Spring Harbor Laboratory,1974609–625.
- 102.
- Cross F, Roberts J, Weintraub H. Simple and complex cell cycles. Annu Rev Cell Biol. 1989;5:341–395. [PubMed: 2574592]
- 103.
- Murray AW, Hunt T. The cell cycle: An introductionNew York: WH Freeman and Co,1993 .
- 104.
- Whitfield JF, Durkin JP, Franks DJ. et al. Calcium, cyclic AMP and protein kinase. Cancer Metast Rev. C—partners in mitogenesis;1987;5:205–250. [PubMed: 3030578]
- 105.
- Dumont JE, Jauniaux J-C, Roger PP. The cyclic AMP-mediated stimulation of cell proliferation. Trends Biochem Sci. 1989;14:67–71. [PubMed: 2539663]
- 106.
- Whitfield JF. Calcium, cell cycles, and cancerBoca Raton, Florida: CRC Press,1990 .
- 107.
- Matsumoto K, Uno I, Ishikawa T. Control of cell division in Saccharomyces cerevisiae mutants defective in adenylate cyclase and cAMP-dependent kinase. Exp Cell Res. 1983;146:151–161. [PubMed: 6305691]
- 108.
- Boynton AL, Whitfield JF. The role of cyclic AMP in cell proliferation: A critical assessment of the evidence. Adv cyclic Nucleotide Res. 1983;15:193–294.
- 109.
- Grieco D, Avvedimento EV, Gotttesman ME. A role for cAMP-dependent protein kinase in early embryonic divisions. Proc Natl Acad Sci USA. 1994;91:9896–9900. [PMC free article: PMC44924] [PubMed: 7937913]
- 110.
- Grieco D, Porcellini A, Avvedimento EV. et al. Requirement for cAMP-PKA pathway activation by M phase-promoting factor in the transition from mitosis to interphase. Science. 1996;271:1718–1723. [PubMed: 8596931]
- 111.
- Carell EF, Dearfield KL. A relationship between adenosine 3'-5'-cyclic monophosphate levels and deoxyribonucleic acid synthesis in Euglena. Life Sci. 1982;31:249–254. [PubMed: 6289032]
- 112.
- Ehsan H, Reichheld J-P, Roef L. et al. Effect of indomethacin on cell cycle dependent cyclic AMP fluxes in tobacco BY-2 cells. FEBS Lett. 1998;442:165–169. [PubMed: 9489998]
- 113.
- Eskin A, Corrent G, Lin C-Y. et al. Mechanism for shifting the phase of a circadian rhythm by serotonin: Involvment of cAMP. Proc Nat Acad Sci USA. 1982;79:660–664. [PMC free article: PMC345806] [PubMed: 6281777]
- 114.
- Prosser RA, Gillette MU. The mammalian circadian clock in the suprachiasmatic nuclei is reset in vitro by cAMP. J Neurosci. 1989;9:1073–1081. [PMC free article: PMC6569955] [PubMed: 2538580]
- 115.
- Zatz M. Does the circadian pacemaker act through cyclic AMP to drive the melatonin rhythm in chick pineal cells? J Biol Rhythms. 1992;7:301–311. [PubMed: 1337482]
- 116.
- Carré IA, Edmunds JrLN. cAMP-dependent kinases in the algal flagellate Euglena gracilis. J Biol Chem. 1992;267:2135–2137. [PubMed: 1310309]
- 117.
- John PCL, Sek FJ, Lee MG. A homolog of the cell cycle control protein p34cdc2 participates in the division cycle of Chlamydomonas, and a similar protein is detectable in higher plants and remote taxa. Plant Cell. 1989;1:1185–1193. [PMC free article: PMC159854] [PubMed: 2535538]
- 118.
- John PCL, Zhang K, Dong C. A p34cdc2-based cell cycle: Its significance in monocotyledonous, dicotyledonous and unicellular plantsIn: Ormond JC, Francis D, eds.Molecular and Cell Biology of the Cell CycleDordrecht: Kluwer Academic Publishers,19939–34.
- 119.
- Bjarnason GA, Jordan RCK, Wood PA. et al. Circadian expression of clock genes in human oral mucosa and skin. Association with specific cell-cycle phases. Am J Pathol. 2001;158:1793–1801. [PMC free article: PMC1891949] [PubMed: 11337377]
- 120.
- Bjarnason GA, Jordan R. Rhythms in human gastrointesdtinal mucosa and skin. Chronobiol Int. 2002;19:129–140. [PubMed: 11962671]
- 121.
- Matsuo T, Yamaguchi S, Mitsui S. et al. Control mechanism of the circadian clock for timing of cell division in vivo. Science. 2003;302:255–259. [PubMed: 12934012]
- 122.
- Granda TG, Liu X-H, Smaaland R. et al. Circadian regulation of cell cycle and apoptosis proteins in mouse bone marrow and tumor FASEB J 200418(101096/fj04-2665fje) [PubMed: 15545298]
- Circadian Organization in the Algal Flagellate Euglena - Madame Curie Bioscience...Circadian Organization in the Algal Flagellate Euglena - Madame Curie Bioscience Database
- Pro-Inflammatory Responses in Macrophages during Toxoplasma gondii Infection - M...Pro-Inflammatory Responses in Macrophages during Toxoplasma gondii Infection - Madame Curie Bioscience Database
- Periodic Selection and Ecological Diversity in Bacteria - Madame Curie Bioscienc...Periodic Selection and Ecological Diversity in Bacteria - Madame Curie Bioscience Database
- Brain Metastasis - Madame Curie Bioscience DatabaseBrain Metastasis - Madame Curie Bioscience Database
- Theory of Organelle Biogenesis: A Historical Perspective - Madame Curie Bioscien...Theory of Organelle Biogenesis: A Historical Perspective - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...