

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

The activities of the Hedgehog (Hh) family of proteins are central to the growth and patterning of developing tissues and organs in many different organisms. Hh proteins are secreted ligands synthesized in discrete regions. The receptor of Hh is Patched (Ptc) and it is expressed in the cells close to the source of Hh. Ptc binds the ligand and transduces a signal which is modulated depending on the context and the concentration of Hh received. Hh and several molecular components of the pathway were first identified and characterized in Drosophila, providing relevant milestones to our understanding on how the Hh signal is transduced. However, important gaps in the pathway still need to be elucidated. Some of these gaps converge on the Ptc receptor and its intriguing mechanisms of Hh reception and signal transduction. Mutations of Ptc that prevail both in animal and human populations are giving some clues on crucial aspects of its function. Patients bearing mutated forms of Ptc suffer a variety of serious diseases. Molecular and cellular studies in Drosophila have given us a clue of the function of Ptc receptor such as the normal topology and/or sorting of the receptor. Thus, a widened knowledge of the function of Ptc might help to design specific therapies for these disorders. This chapter focuses on recent advances that shed some light on how Ptc may operate in the cell.
The Hedgehog Signaling Pathway
Several genes of the Hh pathway were first identified in the fly Drosophila melanogaster and later in vertebrates. Many of the names of the genes involved in the Hh signaling pathway were originally descriptive of the phenotype manifested in mutant Drosophila larvae. Wild-type larvae show a clearly segmented pattern due to alternate bands of denticles in ventral position, whereas the inter-band space is naked. During a screening for mutations that affected the segmental pattern, Nüslein-Volhard and Wieschaus described a group of mutants with alterations in the patterning of the segments.1 Instead of the wild-type alternate belts of denticles and naked cuticle, Hh mutants showed a continuous lawn of denticles, which gave the larva a resemblance to a hedgehog (fig. 1B), and Ptc mutants showed patches of denticles (fig. 1C).
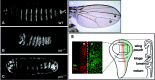
The active Hh protein is a peptide that has undergone an autocatalytical processing in which the peptide is also modified at its N- and C-termini by palmitoyl and cholesterol adducts, respectively. Hh is secreted by discrete subsets of cells within a developing organ in a process that seems mediated, at least in part, by the transmembrane protein Dispatched (Disp), which has a structure very similar to Ptc. In the wing imaginal disc of Drosophila, Hh is expressed in the cells of the posterior compartment and signals to the cells of the anterior compartment, which in turn express the Ptc receptor (fig.1D,E). A graded and short-range response to Hh signaling occurs in the Hh receiving cells (fig. 1E). Ptc, a 12-transmembrane protein, is a negative receptor because it keeps the Hh pathway silenced in its unliganded state.2 In the absence of Hh, Ptc suppresses the activity of Smoothened (Smo), a 7-transmembrane protein that is the positive modulator of the pathway. Although Hh does not bind to Smo, without Smo there is no signaling. The target cells bind Hh by Ptc, which results in the activation of Smo, and, subsequently, in the activation of transcription factors: the Cubitus interruptus protein (Ci) in Drosophila and the orthologous Gli proteins in mammals (reviewed in ref. 10). One peculiarity of the Hh signaling pathway is that Ptc is up-regulated in response to increasing amounts of Hh.3-7 Therefore, Hh controls both its own activity and its own spreading expressing high levels of the receptor.8 By this means, a morphogenetic gradient is formed in the Hh receiving cells. Theoretical analysis of model systems built to explain how a morphogen gradient is formed indicates that this feedback is indeed required to give robustness to the gradient while maintaining its range of action.9 Thus, according to one model, it is expected that fluctuations in ligand production have minor effects on the slope of the gradient, compared to a model in which the receptor has a constant, or ligand-independent level of expression.9
Since this chapter is particularly centered in the Ptc receptor, for a more general view on the Hh response network, the reader is referred to some other recent and exhaustive reviews.10-14
Although this sequence of events is well established, many molecular mechanisms connecting these and other components of the pathway remain elusive. Focusing in the case of the Ptc receptor, several fundamental questions remain unanswered: How does the Ptc receptor recognize the Hh ligand? What happens after the receptor-ligand complex is formed? Is it internalized or does it remain in the membrane before the signal is transduced? How does the unliganded Ptc receptor repress the Smo activity? And how does the liganded Ptc derepress Smo activity?
Sequence and Functional Analysis of Ptc
A single ptc gene is present in Drosophila, whereas two are present in vertebrates (PTCH1 and PTCH2 in humans; Ptc1 and Ptc2 in mice) (reviewed in ref. 10) and several ptc-related genes are found in the nematode worm Caenorhabditis elegans.15 Ptc shows homology to bacterial proton-driven transmembrane molecular transporters,16,17 and presents a conserved a Sterol-Sensing Domain (SSD) that overlaps between transmembrane segment 4 to 6, and also two extracellular large loops (reviewed in ref. 18). In general, SSDs are thought to function as a regulatory domain involved in linking vesicle trafficking and protein localization with processes such as cholesterol homeostasis and cell signaling.18 Concretely, the SSD of Ptc seems to mediate the intracellular trafficking of Ptc, a process that might be essential to regulate Smo activity.19,20 Other functional studies have assigned particular roles to regions of the protein with no obvious similarity to conserved domains. Thus, the two extracellular large loops are required for Hh binding,21 the cytoplasmic C-terminus has been involved in the transduction of the signal,22 and an intracellular small loop between the SSD and the following transmembrane domain has been reported to interact with cyclin B1 to regulate cell-cycle progression in vertebrates.23 However, despite this emerging body of data, we still do not know the full sequence of molecular events that involves the reception of the Hh ligand and how Ptc regulates Smo.
PTCH2 is a 12-transmembrane protein as well and the two large extracellular loops and an SSD are also conserved (reviewed in ref. 24). However, both amino- and carboxy-termini are different from PTCH1. Similar differences are found between Ptc1 and Ptc 2 in mice. PTCH1 and PTCH2 proteins are expressed differentially during the development of the epidermis, suggesting specific roles for each protein,25 although the function of PTCH2 remains unclear to date. A recent study suggests that PTCH2 may act as a Hh receptor that tunes finely the signaling in various cellular environments.26
Hedgehog Lipid Modifications and Morphogen Distribution
The lipid modifications on the Hh protein appear to regulate its activity and distribution. Hedgehog proteins undergo two sequential lipid modifications during their posttranscriptional maturation. Following cleavage of an N-terminus signal sequence upon entry into the secretory pathway, Hh proteins undergo an autoprocessive reaction that results in an internal cleavage inside the 45-KDa precursors, between glycine-cysteine residues from a conserved GCF motif. After this, a cholesterol molecule is covalently added at the newly generated C-terminus of the proteins.27 In Drosophila, a construct of Hh lacking the cholesterol moiety is active in signaling but not appropriately restricted spatially in its signaling activity.28 Therefore, the cholesteryl moiety restricts the spatial deployment of the signal via insertion into the lipid bilayer of the cell membrane and also functions as an essential molecular handle for a proper intracellular and extracellular trafficking and localization of the signal (reviewed in refs. 29, 30). In addition, Hh proteins are palmitoylated on a highly conserved amino-terminal cysteine residue, the first of a pentapeptide CGPGR.31 In Drosophila this palmitoylation is a total requirement in order to produce a fully active Hh signal. Although in vitro studies in vertebrates indicated that Hh mutant forms lacking the pamitoyl adduct retained significant activity,32,33 knockout mice deficient in Skn (the murine ortholog of the Drosophila Ski, which catalyzes Hh palmitoylation) showed that Hh acylation is absolutely required for Hh long-range signaling.24
In summary, dual lipidation of Hh protein promotes membrane affinity and allows its association to sterol-rich membrane microdomains in Drosophila, and to lipid rafts in mammalian cells, that function as platforms for intracellular sorting and signal transduction.34 The close association of Hh proteins to the plasma membrane due to the lipid modifications could provide a mechanism for restricting the range of their activities.
Reception of Hh
Biochemical studies have shown that Hh monomers can form multimeric complexes in which the hydrophobic moieties have been proposed to be sequestered in the interior of the multimer, making the complex soluble and diffusible.35,36 This could be relevant to the movement of Hh through the extracellular matrix37 and to the reception of Hh by Ptc. On one hand, the restricted diffusion of Hh through the extracellular matrix might require the formation of these multimeric complexes of lipidated Hh. On the other hand, a multimeric Hh complex might have either a higher intrinsic affinity for Ptc, or maybe is capable of eliciting greater biological responses, for instance through receptor oligomerization, or binding to coreceptor proteins. In this context, structural studies on transmembrane transporters related to Ptc determined that that proteins operate as homotrimers.38 Interestingly, a mutant version of Ptc that internalizes inefficiently is, however, localized to intracellular vesicles when coexpressed with a dominant negative Ptc.39 This ptc inter-allelic complementation strongly suggests that Ptc has an oligomeric structure.
Several other proteins have been identified as capable of binding to Hh, raising the question of whether they are coreceptors or adjuvants involved in the reception of Hh. Especially, recent analysis in Drosophila have illustrated the critical roles of heparan sulfate proteoglycans (HSPGs) in developmental signaling pathways (reviewed in ref. 40). These large macromolecules are found at the cell surface and form part of the extracellular matrix. A major feature of these proteins is the attachment of long unbranched chains of repeating and sulfated disaccharides to specific serine residues in their protein core. Dally and Dally-like (Dly), two Drosophila glypican members of the HSPG family, has received great attention. A specific role in Hh signaling in the embryonic epidermis has been ascribed to Dly,41 while both Dally and Dly seem to be functionally redundant in the wing disc,42 indicating distinct activities of these two glypicans in the embryo and the wing disc. Hh signaling might also be regulated by other HSPGs in different tissues or developmental processes. For example, Trol, the Drosophila ortholog of Perlecan, seems to form a complex with Hh, and mutations in the gene encoding it cause neuroblasts to undergo cell cycle arrest in the larval brain.43,44 In addition, trol is required for the neuroblast division induced by Hh.43
Two data favour a model in which the polyanionic branches of the HSPGs would act mainly as a molecular trap to keep the morphogens in touch with epithelium surfaces. First, the strongest loss-of-function phenotypes are achieved with mutants affecting the synthesis of sugar chains of the HSPGs;42 and second, the fact that the sugar chains seem to be directly involved in the movement of other morphogens.45,46 Hence, the HSPGs might restrict the morphogen diffusion to the environment of the extracellular matrix, and thus preventing a massive spreading to cavities such as the imaginal disc lumen. This lateral distribution of Hh would facilitate the encounter with Ptc. This scenario does not rule out the possibility of specific regulations involving Dally or Dly. In fact, in tissue culture experiments, Dally-like protein (Dlp), but not Dally was required for Hh signaling.47 In addition, it has been shown that Dlp is specifically required for Hh signaling in the embryonic epidermis.41 A Dlp-mediated regulation of Hh signaling might involve Dlp acting as a coreceptor, perhaps by transferring Hh to Ptc, or by forming a Hh-Dlp-Ptc interaction or stabilizing a Hh-Ptc complex (reviewed in ref. 48). In the case of the Wingless pathway (another morphogenetic signal), it has been elegantly shown that Dlp is cleaved specifically from the cell surface by Notum, a secreted enzyme, resulting in an adequate regulation of the pathway.49
Hh Internalization and Signal Transduction
After receiving Hh, the receptor-ligand complex is internalized and the morphogen is thus depleted from the extracellular milieu. It was proposed that this internalization has a role shaping the Hh gradient.8,50 This hypothesis has been reinforced by experiments that uncoupled the Hh sequestration and the Hh signaling using a Ptc mutant that is unable to sequester Hh and yet retains a normal capacity to mediate Hh signaling.39 Furthermore, the ability to uncouple these functions suggests that Ptc-mediated internalization of Hh does not play a major role in the transduction of the signal but serves mainly to limit and control the Hh gradient.39
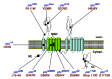
The analysis of different ptc mutants (fig. 2) in Drosophila have demonstrated that the two functions of Ptc, sequestration and signalling, can be genetically separated.8,19,20,22,39,51 PtcS2, Ptcm13, Ptc34 and Ptc1130 are defective for Hh signal transduction but sequestration properties are normal,19,20,22 while the Ptc14 mutant protein is defective for Hh sequestration, but normal in terms of Hh signal transduction.39
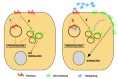
One of the most intriguing aspects of the Hh pathway is how Ptc regulates Smo activity. The mechanism is most probably indirect, since Ptc and Smo do not need to bind or to colocalize to control signaling and, in addition, Ptc acts substoichometrically to inhibit Smo, possibly through changes in the distribution or concentration of an as yet uncharacterized small molecule (fig. 3).52 A recent study53 suggests a mechanism by which the ratio of unliganded to liganded Ptc determines the cellular response (reviewed in ref. 54). In this context, several alternative models are possible. For example, unliganded Ptc might import a Smo antagonist, while liganded Ptc might promote its export. Alternatively, the functional Hh receptor may comprise a multimer of Ptc proteins and the binding of Hh to one Ptc subunit in a receptor complex may block the ability of the multimer to inhibit Smo activitity.53,54
Smo Regulation by Ptc
When Hh binds Ptc, Smo is somehow liberated to start the process of activating target gene transcription. The mechanism in which Ptc inhibits and Hh activates Smo protein remains unknown. Smo is transcribed in a generalized pattern not transcriptionally regulated by the Hh signal. Smo protein is posttranscriptionally regulated by phosphorylation, becoming more stable, and by moving to the cell surface after a Hh signal is received.55-60 A detailed study has shown that Smo accumulates in the plasma membrane of cells in which Ptc activity is abrogated by Hh but is targeted to the degradative pathway in cells where Ptc is active.61 Thus, Ptc could regulate Smo activity through inhibition of its accumulation in the plasma membrane, targeting instead to lysosomal compartments. In this view, Ptc might regulate Smo activity simply by modulating the levels of protein present in the cell. Alternatively, it may be that sub-cellular location of Smo is critical for its activation, the plasma membrane providing an environment for Smo to be accessible to an intracellular agonist (see fig. 3 for details).61
Ptc and Human Disease
Genetic studies in various model organisms are beginning to elucidate the factors that are likely candidates for the causes of early embryonic defects in humans (reviewed in refs. 62, 63). Thus, detailed knowledge of the Hh signaling pathway is fundamental to an understanding of vertebrate development as well as several birth defects in humans.
Since the unliganded Ptc receptor exerts a repression on the Hh pathway, mutations affecting Ptc are frequently associated to a constitutive activation of the Hh signaling pathway. This is in agreement with the fact that most of the mutations characterized to date in PTCH1 result in protein truncation.24 The spectrum of human PTCH1 mutations also includes deletions, insertions, splice site alterations, and nonsense and missense mutations distributed throughout the gene (for an example, see Table 1).64-66
Table 1
Mutations of the PTCH1 gene detected in patients with Gorlin Syndrome and BCC (see refs. ).
PTCH1 deregulation in the epidermis is sufficient to induce Basal Cell Carcinomas (BCCs) of the skin. 30-40% of Gorlin syndrome patients (GS; also known as Nevoid basal cell carcinoma syndrome) have familial loss-of-function mutations in the PTCH1 gene. GS is an autosomal dominant disease with nearly complete penetrance and variable expressivity because mutations in PTCH1 seem to be haploinsuficient in humans (a mutation in only one of the two alleles is enough to show a phenotype). Clinically, GS patients present congenital abnormalities that includes skeletal defects (polydactyly, fused or bifid ribs), early onset of multiple BCCs and an increased rate to develop other tumors, including medulloblastomas of the cerebellum (reviewed in refs. 67-69). A limited number of mutations in PTCH1 have been linked to Holoprosencephaly (HPE).70 HPE affects the forebrain and face to various degrees, from the most extreme lethal alobar type to milder microforms that include small midline facial defects. However, since HPE arises from loss of SHH signaling, these mutations might rather reflect an impaired ability of PTCH1 to interact with SHH, thus permanently shutting down the pathway. In fact, two out of four rare PTCH1 missense mutations that have been reported to be associated to HPE were localized in the extracellular loops of PTCH1 required for SHH binding.70
There is a limited number of missense mutations described for PTCH1. Missense mutations do not occur as frequently as frameshift or nonsense mutations. In a clinical study on French patients, missense mutations span the second group of six transmembrane domains of the protein.64 Unfortunately, no data are available to interpret the pathologic value of the missense mutations described in humans.64 So far, only the molecular and cellular studies of Drosophila ptc mutations indicate an alteration of the correct topology and/or sorting of the Ptc receptor. We propose Drosophila as an ideal model system to analyze the functional alterations of the missense mutations found in the PTCH1 receptor. In fact, a recent study analyses in the fly the function of an artificially mutated form of Ptc analog to a mutated PTCH1 previously characterized in GS patients.71 Therefore, understanding the molecular mechanisms of Hh signal reception will allow identifying potential drug targets in order to devise strategies for the treatment of BCC.
Concluding Remarks
More genes of the Hh pathway are going to be uncovered in the near future and Drosophila genetics will undoubtedly help in to resolve this issue. Hence, concerning the function of Ptc receptor, an unresolved question is how Ptc is able to repress the activity of Smo, and whether there are intermediate elements. Although other molecules have been found as possible candidates in the reception of Hh in vertebrates such as Megalin, Hip or Gas1, mutagenesis screens need to be done in Drosophila to find more proteins implicated in Hh reception. The study of their function will help to understand how several cellular and molecular processes drive and regulate the pathway. Another important unresolved issue in the Hh pathway is the role of cell polarity in the transduction of the signal, or the endocytic and exocytic routes involved. Also questions remain regarding how a morphogen gradient is formed, how the cytoskeleton affects the pathway, or how the transcription of target genes is regulated. New model systems in vertebrate genetics, cellular biology and biochemistry will help to elucidate some of these questions. Definitely, a huge biochemical and cell biology effort needs to be done in order to solve the role of this kind of proteins in Hh signaling pathway. This knowledge will undoubtedly help to design specific therapies to serious human diseases caused by mutations on genes of this signaling pathway.
Acknowledgments
We thank Cristina Guerra-Giraldez for critical reading the manuscript. LQ is supported by an I3P contract of the CSIC; AC is supported by Comunidad Autónoma de Madrid, grants (08.6/0045/2001.2 and GR/SAL/0185/2004); and CT was supported by by the Spanish D.G.I.C.Y.T, grant (BMC2002-03839). Also, an institutional grant from Fundación Ramón Areces is acknowledged.
References
- 1.
- Nusslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980;287(5785):795–801. [PubMed: 6776413]
- 2.
- Ingham PW, Taylor AM, Nakano Y. Role of the Drosophila patched gene in positional signalling. Nature. 1991;353(6340):184–187. [PubMed: 1653906]
- 3.
- Marigo V, Tabin CJ. Regulation of patched by sonic hedgehog in the developing neural tube. Proc Natl Acad Sci USA. 1996;93(18):9346–9351. [PMC free article: PMC38430] [PubMed: 8790332]
- 4.
- Goodrich LV, Johnson RL, Milenkovic L. et al. Conservation of the hedgehog/patched signaling pathway from flies to mice: Induction of a mouse patched gene by Hedgehog. Genes Dev. 1996;10(3):301–312. [PubMed: 8595881]
- 5.
- Capdevila J, Pariente F, Sampedro J. et al. Subcellular localization of the segment polarity protein patched suggests an interaction with the wingless reception complex in Drosophila embryos. Development. 1994;120(4):987–998. [PubMed: 7600973]
- 6.
- Ingham PW, Fietz MJ. Quantitative effects of hedgehog and decapentaplegic activity on the patterning of the Drosophila wing. Curr Biol. 1995;5(4):432–440. [PubMed: 7627558]
- 7.
- Tabata T, Kornberg TB. Hedgehog is a signaling protein with a key role in patterning Drosophila imaginal discs. Cell. 1994;76(1):89–102. [PubMed: 8287482]
- 8.
- Chen Y, Struhl G. Dual roles for patched in sequestering and transducing Hedgehog. Cell. 1996;87(3):553–563. [PubMed: 8898207]
- 9.
- Eldar A, Rosin D, Shilo BZ. et al. Self-enhanced ligand degradation underlies robustness of morphogen gradients. Dev Cell. 2003;5(4):635–646. [PubMed: 14536064]
- 10.
- Ingham PW, McMahon AP. Hedgehog signaling in animal development: Paradigms and principles. Genes Dev. 2001;15(23):3059–3087. [PubMed: 11731473]
- 11.
- Nybakken K, Perrimon N. Hedgehog signal transduction: Recent findings. Curr Opin Genet Dev. 2002;12(5):503–511. [PubMed: 12200154]
- 12.
- Lum L, Beachy PA. The hedgehog response network: Sensors, switches, and routers. Science. 2004;304(5678):1755–1759. [PubMed: 15205520]
- 13.
- Ogden SK, Ascano JrM, Stegman MA. et al. Regulation of Hedgehog signaling: A complex story. Biochem Pharmacol. 2004;67(5):805–814. [PMC free article: PMC3659400] [PubMed: 15104233]
- 14.
- Bijlsma MF, Spek CA, Peppelenbosch MP. Hedgehog: An unusual signal transducer. Bioessays. 2004;26(4):387–394. [PubMed: 15057936]
- 15.
- Aspock G, Kagoshima H, Niklaus G. et al. Caenorhabditis elegans has scores of hedgehog-related genes: Sequence and expression analysis. Genome Res. 1999;9(10):909–923. [PubMed: 10523520]
- 16.
- Nakano Y, Guerrero I, Hidalgo A. et al. A protein with several possible membrane-spanning domains encoded by the Drosophila segment polarity gene patched. Nature. 1989;341(6242):508–513. [PubMed: 2797178]
- 17.
- Hooper JE, Scott MP. The Drosophila patched gene encodes a putative membrane protein required for segmental patterning. Cell. 1989;59(4):751–765. [PubMed: 2582494]
- 18.
- Kuwabara PE, Labouesse M. The sterol-sensing domain: Multiple families, a unique role? Trends Genet. 2002;18(4):193–201. [PubMed: 11932020]
- 19.
- Martin V, Carrillo G, Torroja C. et al. The sterol-sensing domain of Patched protein seems to control Smoothened activity through Patched vesicular trafficking. Curr Biol. 2001;11(8):601–607. [PubMed: 11369205]
- 20.
- Strutt H, Thomas C, Nakano Y. et al. Mutations in the sterol-sensing domain of Patched suggest a role for vesicular trafficking in Smoothened regulation. Curr Biol. 2001;11(8):608–613. [PubMed: 11369206]
- 21.
- Briscoe J, Chen Y, Jessell TM. et al. A hedgehog-insensitive form of patched provides evidence for direct long-range morphogen activity of sonic hedgehog in the neural tube. Mol Cell. 2001;7(6):1279–1291. [PubMed: 11430830]
- 22.
- Johnson RL, Milenkovic L, Scott MP. In vivo functions of the patched protein: Requirement of the C terminus for target gene inactivation but not Hedgehog sequestration. Mol Cell. 2000;6(2):467–478. [PubMed: 10983992]
- 23.
- Barnes EA, Kong M, Ollendorff V. et al. Patched1 interacts with cyclin B1 to regulate cell cycle progression. Embo J. 2001;20(9):2214–2223. [PMC free article: PMC125436] [PubMed: 11331587]
- 24.
- Cohen JrMM. The hedgehog signaling network. Am J Med Genet A. 2003;123(1):5–28. [PubMed: 14556242]
- 25.
- Smyth I, Narang MA, Evans T. et al. Isolation and characterization of human patched 2 (PTCH2), a putative tumour suppressor gene inbasal cell carcinoma and medulloblastoma on chromosome 1p32. Hum Mol Genet. 1999;8(2):291–297. [PubMed: 9931336]
- 26.
- Rahnama F, Toftgard R, Zaphiropoulos PG. Distinct roles of PTCH2 splice variants in Hedgehog signalling. Biochem J. 2004;378(Pt 2):325–334. [PMC free article: PMC1223965] [PubMed: 14613484]
- 27.
- Porter JA, Young KE, Beachy PA. Cholesterol modification of hedgehog signaling proteins in animal development. Science. 1996;274(5285):255–259. [PubMed: 8824192]
- 28.
- Porter JA, Ekker SC, Park WJ. et al. Hedgehog patterning activity: Role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell. 1996;86(1):21–34. [PubMed: 8689684]
- 29.
- Mann RK, Beachy PA. Novel lipid modifications of secreted protein signals. Annu Rev Biochem. 2004;73:891–923. [PubMed: 15189162]
- 30.
- Mann RK, Beachy PA. Cholesterol modification of proteins. Biochim Biophys Acta. 2000;1529(1-3):188–202. [PubMed: 11111088]
- 31.
- Pepinsky RB, Zeng C, Wen D. et al. Identification of a palmitic acid-modified form of human Sonic hedgehog. J Biol Chem. 1998;273(22):14037–14045. [PubMed: 9593755]
- 32.
- Lee JD, Kraus P, Gaiano N. et al. An acylatable residue of Hedgehog is differentially required in Drosophila and mouse limb development. Dev Biol. 2001;233(1):122–136. [PubMed: 11319862]
- 33.
- Kohtz JD, Lee HY, Gaiano N. et al. N-terminal fatty-acylation of sonic hedgehog enhances the induction of rodent ventral forebrain neurons. Development. 2001;128(12):2351–2363. [PubMed: 11493554]
- 34.
- Rietveld A, Neutz S, Simons K. et al. Association of sterol- and glycosylphosphatidylinositol-linked proteins with Drosophila raft lipid microdomains. J Biol Chem. 1999;274(17):12049–12054. [PubMed: 10207028]
- 35.
- Chen MH, Li YJ, Kawakami T. et al. Palmitoylation is required for the production of a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. Genes Dev. 2004;18(6):641–659. [PMC free article: PMC387240] [PubMed: 15075292]
- 36.
- Zeng X, Goetz JA, Suber LM. et al. A freely diffusible form of Sonic hedgehog mediates long-range signalling. Nature. 2001;411(6838):716–720. [PubMed: 11395778]
- 37.
- The I, Bellaiche Y, Perrimon N. Hedgehog movement is regulated through tout velu-dependent synthesis of a heparan sulfate proteoglycan. Mol Cell. 1999;4(4):633–639. [PubMed: 10549295]
- 38.
- Murakami S, Nakashima R, Yamashita E. et al. Crystal structure of bacterial multidrug efflux transporter AcrB. Nature. 2002;419(6907):587–593. [PubMed: 12374972]
- 39.
- Torroja C, Gorfinkiel N, Guerrero I. Patched controls the Hedgehog gradient by endocytosis in a dynamin-dependent manner, but this internalization does not play a major role in signal transduction. Development. 2004;131(10):2395–2408. [PubMed: 15102702]
- 40.
- Baeg GH, Perrimon N. Functional binding of secreted molecules to heparan sulfate proteoglycans in Drosophila. Curr Opin Cell Biol. 2000;12(5):575–580. [PubMed: 10978892]
- 41.
- Desbordes SC, Sanson B. The glypican Dally-like is required for Hedgehog signalling in the embryonic epidermis of Drosophila. Development. 2003;130(25):6245–6255. [PubMed: 14602684]
- 42.
- Han C, Belenkaya TY, Wang B. et al. Drosophila glypicans control the cell-to-cell movement of Hedgehog by a dynamin-independent process. Development. 2004;131(3):601–611. [PubMed: 14729575]
- 43.
- Park Y, Rangel C, Reynolds MM. et al. Drosophila perlecan modulates FGF and hedgehog signals to activate neural stem cell division. Dev Biol. 2003;253(2):247–257. [PubMed: 12645928]
- 44.
- Voigt A, Pflanz R, Schafer U. et al. Perlecan participates in proliferation activation of quiescent Drosophila neuroblasts. Dev Dyn. 2002;224(4):403–412. [PubMed: 12203732]
- 45.
- Han C, Belenkaya TY, Khodoun M. et al. Distinct and collaborative roles of Drosophila EXT family proteins in morphogen signalling and gradient formation. Development. 2004;131(7):1563–1575. [PubMed: 14998928]
- 46.
- Takei Y, Ozawa Y, Sato M. et al. Three Drosophila EXT genes shape morphogen gradients through synthesis of heparan sulfate proteoglycans. Development. 2004;131(1):73–82. [PubMed: 14645127]
- 47.
- Lum L, Yao S, Mozer B. et al. Identification of Hedgehog pathway components by RNAi in Drosophila cultured cells. Science. 2003;299(5615):2039–2045. [PubMed: 12663920]
- 48.
- Lin X. Functions of heparan sulfate proteoglycans in cell signaling during development. Development. 2004;131(24):6009–6021. [PubMed: 15563523]
- 49.
- Kreuger J, Perez L, Giraldez AJ. et al. Opposing activities of dally-like glypican at high and low levels of wingless morphogen activity. Dev Cell. 2004;7(4):503–512. [PubMed: 15469839]
- 50.
- Gallet A, Therond PP. Temporal modulation of the Hedgehog morphogen gradient by a patched-dependent targeting to lysosomal compartment. Dev Biol. 2005;277(1):51–62. [PubMed: 15572139]
- 51.
- Vegh M, Basler K. A genetic screen for hedgehog targets involved in the maintenance of the Drosophila anteroposterior compartment boundary. Genetics. 2003;163(4):1427–1438. [PMC free article: PMC1462513] [PubMed: 12702686]
- 52.
- Taipale J, Cooper MK, Maiti T. et al. Patched acts catalytically to suppress the activity of Smoothened. Nature. 2002;418(6900):892–897. [PubMed: 12192414]
- 53.
- Casali A, Struhl G. Reading the Hedgehog morphogen gradient by measuring the ratio of bound to unbound Patched protein. Nature. 2004;431(7004):76–80. [PubMed: 15300262]
- 54.
- Briscoe J. Hedgehog signaling: Measuring ligand concentrations with receptor ratios. Curr Biol. 2004;14(20):R889–891. [PubMed: 15498480]
- 55.
- Denef N, Neubuser D, Perez L. et al. Hedgehog induces opposite changes in turnover and subcellular localization of patched and smoothened. Cell. 2000;102(4):521–531. [PubMed: 10966113]
- 56.
- Alcedo J, Zou Y, Noll M. Posttranscriptional regulation of smoothened is part of a self-correcting mechanism in the Hedgehog signaling system. Mol Cell. 2000;6(2):457–465. [PubMed: 10983991]
- 57.
- Ingham PW, Nystedt S, Nakano Y. et al. Patched represses the Hedgehog signalling pathway by promoting modification of the Smoothened protein. Curr Biol. 2000;10(20):1315–1318. [PubMed: 11069117]
- 58.
- Zhu AJ, Zheng L, Suyama K. et al. Altered localization of Drosophila Smoothened protein activates Hedgehog signal transduction. Genes Dev. 2003;17(10):1240–1252. [PMC free article: PMC196058] [PubMed: 12730121]
- 59.
- Zhang C, Williams EH, Guo Y. et al. Extensive phosphorylation of Smoothened in Hedgehog pathway activation. Proc Natl Acad Sci USA. 2004 [PMC free article: PMC535705] [PubMed: 15598741]
- 60.
- Jia J, Tong C, Wang B. et al. Hedgehog signalling activity of Smoothened requires phosphorylation by protein kinase A and casein kinase I. Nature. 2004;432(7020):1045–1050. [PubMed: 15616566]
- 61.
- Nakano Y, Nystedt S, Shivdasani AA. et al. Functional domains and sub-cellular distribution of the Hedgehog transducing protein Smoothened in Drosophila. Mech Dev. 2004;121(6):507–518. [PubMed: 15172682]
- 62.
- Mullor JL, Sanchez P, Ruiz i Altaba A. Pathways and consequences: Hedgehog signaling in human disease. Trends Cell Biol. 2002;12(12):562–569. [PubMed: 12495844]
- 63.
- Roessler E, Muenke M. How a Hedgehog might see holoprosencephaly. Hum Mol Genet. 2003;12(Spec No 1):R15–25. [PubMed: 12668593]
- 64.
- Boutet N, Bignon YJ, Drouin-Garraud V. et al. Spectrum of PTCH1 mutations in French patients with Gorlin syndrome. J Invest Dermatol. 2003;121(3):478–481. [PubMed: 12925203]
- 65.
- Chidambaram A, Goldstein AM, Gailani MR. et al. Mutations in the human homologue of the Drosophila patched gene in Caucasian and African-American nevoid basal cell carcinoma syndrome patients. Cancer Res. 1996;56(20):4599–4601. [PubMed: 8840969]
- 66.
- Hahn H, Wojnowski L, Miller G. et al. The patched signaling pathway in tumorigenesis and development: Lessons from animal models. J Mol Med. 1999;77(6):459–468. [PubMed: 10475061]
- 67.
- Ruiz i Altaba A, Sanchez P, Dahmane N. Gli and hedgehog in cancer: Tumours, embryos and stem cells. Nat Rev Cancer. 2002;2(5):361–372. [PubMed: 12044012]
- 68.
- Goodrich LV, Scott MP. Hedgehog and patched in neural development and disease. Neuron. 1998;21(6):1243–1257. [PubMed: 9883719]
- 69.
- Taipale J, Beachy PA. The Hedgehog and Wnt signalling pathways in cancer. Nature. 2001;411(6835):349–354. [PubMed: 11357142]
- 70.
- Ming JE, Kaupas ME, Roessler E. et al. Mutations in PATCHED-1, the receptor for SONIC HEDGEHOG, are associated with holoprosencephaly. Hum Genet. 2002;110(4):297–301. [PubMed: 11941477]
- 71.
- Hime GR, Lada H, Fietz MJ. et al. Functional analysis in Drosophila indicates that the NBCCS/ PTCH1 mutation G509V results in activation of smoothened through a dominant-negative mechanism. Dev Dyn. 2004;229(4):780–790. [PubMed: 15042702]
- 72.
- Mullor JL, Guerrero I. A gain-of-function mutant of patched dissects different responses to the hedgehog gradient. Dev Biol. 2000;228(2):211–224. [PubMed: 11112325]
- 73.
- Capdevila J, Estrada MP, Sanchez-Herrero E. et al. The Drosophila segment polarity gene patched interacts with decapentaplegic in wing development. Embo J. 1994;13(1):71–82. [PMC free article: PMC394780] [PubMed: 8306973]
- 74.
- Strange RC, El-Genidy N, Ramachandran S. et al. Susceptibility to basal cell carcinoma: Associations with PTCH Polymorphisms. Ann Hum Genet. 2004;68(Pt 6):536–545. [PubMed: 15598212]
- The Patched Receptor: Switching On/Off the Hedgehog Signaling Pathway - Madame C...The Patched Receptor: Switching On/Off the Hedgehog Signaling Pathway - Madame Curie Bioscience Database
- RNA NETWORKS IN PROKARYOTES II: tRNA Processing and Small RNAs - Madame Curie Bi...RNA NETWORKS IN PROKARYOTES II: tRNA Processing and Small RNAs - Madame Curie Bioscience Database
- DNA Packaging by Bacteriophage P22 - Madame Curie Bioscience DatabaseDNA Packaging by Bacteriophage P22 - Madame Curie Bioscience Database
- Regulation of Insulin Action and Insulin Secretion by SNARE-Mediated Vesicle Exo...Regulation of Insulin Action and Insulin Secretion by SNARE-Mediated Vesicle Exocytosis - Madame Curie Bioscience Database
- Structure, Function and Assembly of Flagellar Axial Proteins - Madame Curie Bios...Structure, Function and Assembly of Flagellar Axial Proteins - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...