

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Zebrafish is now a well established model for the study of developmental and regenerative processes. Indeed, the genetic cascades that control the early development of the structure that will form the paired fins (the fin bud) present similarities with the early formation of the tetrapod fore and hindlimb buds. One of these conserved molecular pathways involves secreted factors of the Hedgehog family [sonic hedgehog (shh) and tiggywinkle hedgehog (twhh)]. As in the tetrapod limbs, hedgehog proteins are initially expressed in the posterior region of the early fin bud where they contribute to the patterning of the antero-posterior axis, then are involved in cell proliferation and the formation of various skeletal elements. The hedgehog pathway is reactivated in adult fish following fin amputation, an event that triggers the regeneration program. During this process, the hedgehog signal is involved in various processes such as the growth and maintenance of the blastema and patterning of the fin ray.
The Zebrafish Hedgehog Genes
Vertebrate Hedgehog (Hh) genes are classified into three classes: sonic (shh), indian (ihha) and desert (dhh*) hedgehog class. Most vertebrate species possess one member from each gene family. However, the teleost danio rerio (zebrafish) possesses at least five hedgehog genes: two sonic-class genes: shh1 and tiggywinkle (twhh),2 two Indian-class genes: echidna (ehh)3 and ihh4 and one desert-class gene4 (fig. 1). Despite the high number of Hh genes, we will see that only shh and twhh are expressed during fin bud development and only shh seems to be required for their proper development. During fin ray regeneration, both shh and the newly identified ihh are expressed, and functional data indicates that the Hh signaling pathway is involved in blastema formation and maintenance, and later in fin ray patterning.
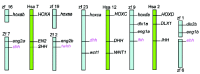
Figure 1
Chromosomal location of human (Hsa) and zebrafish (zf ) Hedgehog genes.
Overview of the Zebrafish Pectoral Fin Bud Development
Zebrafish possesses five sets of fins divided into two types (fig. 2A,B): the paired fins (pectoral and pelvic) and the median fins (dorsal, caudal and anal). The development of the two types of fins is somehow different: the median fins develop directly from the epidermal fold surrounding the caudal half of the young larvae (the median fin fold5), whereas the paired fins first arise from a local proliferation of the lateral plate mesoderm to form the fin bud.6 However, the visible part of the pectoral fins which contains the exoskeleton (the fin rays) eventually develops inside of an epidermal fin fold in a process that resembles the development of the median fins from the median fin fold.5 Early fish fin buds and tetrapod limb buds show striking morphological resemblances and they both contain equivalent signaling centers: The ZPA (zone of polarizing activity) in the posterior mesenchyme,1,7,8 the apical ectoderm6,9,10 (equivalent to the apical ectodermal ridge, or AER, in tetrapods), and the ventral ectoderm,11,12 which are responsible for the specification of the antero-posterior, proximo-distal and dorso-ventral axes, respectively. However, the AER of the tetrapod limb progressively degenerates during development, whereas the zebrafish apical ectoderm will form an elongated fin fold in which the external part of the fin, including the fin rays will eventually develop.6 The divergence of the outcome of the apical epidermis between tetrapod limb and larval fin is thought to be a major component of the initial morphological differences between the two types of appendages.

One of the molecular pathways involved in both limb and fin development is the hedgehog (Hh) signaling pathway. This pathway has been extensively studied in zebrafish, in particular in the pectoral fin bud which constitutes a practical and accessible model for developmental and functional studies of early limb development.
Fin Bud and Early Larval Fin
The early pectoral fin buds arise by 24 hours-post-fertilization (hpf ) from the limb fields which consist of a pair of small aggregates of mesenchymal cells located on each side of the main body axis at the level of the third somite. As the fin buds grow, the first skeletal elements start to condensate by 37hpf in the center of the fin bud, and will give rise few hours later to the cartilaginous endochondral disk.6 This chondrogenic condensation divides the mesenchymal cell population into a ventral and a dorsal half which will give rise to the muscles of the fin. The proximal part of the chondrogenic condensation will differentiate into the larval endoskeletal girdle and the distal part will develop as the endochondral disc which will give rise to the fin endoskeleton. At 28hpf, the bud is covered by a two-layered epidermis composed of one basal stratum and one flat peridermal cell layer. At about 31hpf, the apical epidermal cells lining the anterior-posterior axis of the bud thicken to form a transient ridge which is similar to the AER of the tetrapod limbs. By 34hpf, the apical epidermal cells undergo a morphological change, detach from the underlying mesenchyme and progressively form an epidermal fold separated by a subepidermal space.1,6 At 48hpf, this epidermal fold starts to elongate and mesenchymal cells start to invade the structure. The actinotrichia, collagenous fibers, are the first supportive elements to form within the larval fin fold, as no fin rays have yet appeared at that stage.
Adult Fin Formation
During the third week of development, the larval fin undergoes massive rearrangement of the endoskeleton and musculature, as it switches from larval to adult shape. Following the rotation of the larval fin, drastic remodeling of the endoskeleton occurs. Then, specialized cells from the distal mesenchyme, the scleroblasts, intercalate between the actinotrichia and the basement membrane and start to secrete the bone matrix in a proximal to distal fashion, forming the fin rays or lepidotrichia. This type of bone (called dermal or intramembranous) is directly mineralized in the subepidermal space, unlike the endochondral bone which is formed through a cartilaginous precursor. Fin ray structure and morphogenesis are further described below (fig. 2B, fig. 5A).

Shh and Twhh Expression during Fin Bud Development
The first sign of shh expression appears by 26-28hpf, in very few cells of the posterior region of the early pectoral fin bud.1,7 By 30hpf, shh expression intensifies and is now clearly restricted to the posterior margin of the fin bud, in a position which is considered to be analogous to the zone of polarizing activity (ZPA) of the tetrapod limb bud (see next paragraph). By 48hpf, shh is expressed in a wider domain but limited to the posterior part of the fin bud. This expression is maintained until approximately 3 day-post-fertilization (dpf) (fig. 2C,D), and is then progressively downregulated until shh transcripts become undetectable by in situ hybridization by 4dpf.13-15 twhh and shh have very similar expression patterns in structures of the embryonic axis such as the notochord, the floor plate and the branchial arches.2,16 However, twhh expression is not detected as early as shh in the fin bud but starts at 48hpf in a group of cells corresponding to shh-expressing cells.6,17 During morphogenesis of the rays of all fin types which initiates around the fourth week post-fertilization, shh is reexpressed in the basal epidermal layer (BEL) at the tip of each forming lepidotrichia (fig. 2E), in a pattern reminiscent to that observed during fin regeneration (see below).15
Shh, Retinoic Acid Regulation and the ZPA
The ZPA is a signaling center located in the posterior mesenchyme of the tetrapod limb and is responsible for the patterning of the anteroposterior (A/P) axis of the limb.18 Shh and retinoic acid are two signaling molecules involved in this process.1,7,8,19-22 In chick embryos, grafting experiments of cells of the ZPA as well as ectopic expression of shh or local application of retinoic acid (RA) at the anterior margin of the limb bud result in the formation of a mirror-image duplication of the digits,8,22,23 thus connecting the polarizing activity of this region to the role of shh and RA. In zebrafish, retinoic acid treatment of embryos at 24-30hpf causes the formation of an ectopic shh expression domain in the anterior region of the pectoral fin bud between 2-3dpf,7,14 and later (by 4dpf), signs of a duplication of the fin bud have been observed, reminiscent of the digit duplication observed in chick.13 It is interesting to note that although RA was delivered to whole zebrafish embryos in these experiments, while locally applied to the chick limb buds,22 both treatments lead to a similar ectopic expression of shh and limb/fin structure duplication. This suggests that very few cells have the potential to form a polarizing zone. Hoffman et al observed that a two hour RA treatment of 30hpf zebrafish embryos causes a transient downregulation of shh, followed by the reappearance, a few hours later, of the posterior shh domain.14 This domain progressively extends towards the anterior region of the bud. Once shh is activated in cells at the anterior margin, its expression is down-regulated in cells of the center of the bud, therefore leaving, 24 hours after the end of the treatment, two discrete anterior and posterior shh domains in the bud. Similar RA treatments of mutants of the shh gene (syu, see next paragraph) lead to the same result indicating that this anterior ectopic expression of shh is independent of shh signaling but could depend on factors secreted by the AER. hoxd-11 and hoxd-12 which are normally expressed in the posterior mesenchyme of the fin bud, present an anterior extension of their expression domain after early RA treatment (at 5hpf) of wild-type embryos, but not of syu mutant embryos.24 Thus, the anterior expansion of these genes by RA is independent of shh signaling suggesting that hox genes could represent of the intermediate factors between shh and RA.14,24 Surprisingly, a DR5-type retinoic acid receptor binding sequence (or retinoic acid response element, RARE) has been identified in the promoter region of the shh gene in zebrafish only.25 This element is functional in vitro, and could directly link retinoic acid to shh. However, no mutation experiment has brought clear evidence about the activity of this element in vivo, and further investigation will clarify its potential function during zebrafish fin bud development.
Mutants of the Hh Pathway and Fin Bud Development
Shh function in zebrafish has been studied using a group of mutants presenting somite formation defects (the you-type mutants, see Table 1), including the syu mutant in which the shh gene is disrupted26 (Table 2). All the mutated genes of the you-type family identified so far are involved in the Hh signaling pathway. Interestingly, mutation of individual genes of the Hh pathway in zebrafish leads to relatively mild and variable phenotypes that are not lethal before several days of development. Two mutants, syu and smu present defects of fin bud development26-28 providing a valuable tool to study the function of the Hh pathway in the development and morphogenesis of the zebrafish fins.
Table 1
Zebrafish you-type genes and mutants.
Table 2
Alleles of the zebrafish syu mutants.
The most obvious phenotypes of the syu (and smu) mutants are defects of the embryonic axis: the embryos show a strong body curvature, U-shaped somites, underdeveloped eyes and jaw-related structures. In addition to these defects, the syu embryos present a wide range of allele-dependent alterations of pectoral fin development.26,29 The two weak alleles (syutq252,syutqb70) provoke moderate and variable reduction of the fin fold and the fin endoskeleton, whereas the strong allele syutbx392 causes a drastic reduction of both the fin bud and fin fold. Embryos homozygous for the deletion allele syut4 initiate fin bud development, but fin growth is not sustained and the mutant completely lack pectoral fins (Table 2 and fig. 3). At the molecular and cellular levels the disruption of shh activity in the early fin bud has three major consequences: disorganization of the A/P patterning, failure to develop and maintain a proper distal epidermis and a decrease in cell proliferation. The expression of the posterior hox genes, which are involved in the A/P patterning, is perturbed in syu embryos. For example, hoxd-13 expression seems to be completely dependent on shh activity as this gene fails to be expressed in syu mutants. In contrast, hoxd-11 and hoxd-12 transcription initiates in a shh-independent fashion but requires the shh signal to be maintained in the posterior region of the fin bud and hoxd-10 expression seems totally independent of shh signaling.24 This raises the question of the factors on which these genes rely to initiate their expression.b Twhh is unlikely to be one of these factors, as its expression is activated later than shh.17 RA has been shown to induce shh expression, even in the absence of the shh signal, showing that it is an important factor for the early specification of the fin bud and could be one of these molecules.
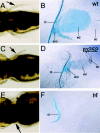
Another phenotype of the syu mutant fin is the shortening or the absence of the pectoral fin fold (fig. 3, left panel), and in the early syut4 the total absence of a normal apical ectoderm (AE). Phenotypically, the early AE forms normally in syu mutants carrying the moderate or the weak allele, but degenerates a few hours later. Expression of the transcription factor dlx2 or the secreted factor fgf2 normally initiates during the early formation of the AE in syu mutants, but it is quickly downregulated at the stage corresponding to fin fold elongation in wild type embryos.24 These observations show that shh activity is not necessary to initiate the expression of distal markers or to specify the distal epidermis, but it is required to maintain the AE integrity and the expression of the distal markers.24 For instance, fgf8, a late marker of the AE, whose expression depends on fgf2 expression, completely fails to be expressed in the syu mutants.24,30 In addition to the problems caused to the A/P patterning and the formation of the distal epidermis, shh disruption causes a decrease in cell proliferation throughout the whole fin bud that leads to defects in cartilage condensation and bone formation (fig. 3, right panel). The decrease in cell proliferation is detected before the formation of the AE, suggesting that the proliferation problems in syu mutants are initially not due to the absence of factors secreted by the distal epidermis. However, the subsequent downregulation of the factors expressed in the AE is likely to aggravate the proliferation defect.
Fin Ray Morphogenesis and Regeneration
The fin rays or lepidotrichia are the skeletal elements of the external part of the adult fins (fig. 2B) which develop relatively late, during the fourth week of life.15,31 The base of the lepidotrichia is attached to the fin endoskeleton via muscles and ligaments. The lepidotrichia are composed of two hemirays, shaped like parenthesis and facing each other (fig. 4A). They are segmented and periodically bifurcate along the proximodistal axis. Each segment is attached to the next one by a collagenous ligament, forming a joint that gives flexibility to the fin ray. Blood vessels, nerves, pigment cells and connective tissue are located between the two hemirays and also in the inter-ray region.

The lepidotrichia are formed in two steps: first, the bone matrix is secreted in the subepidermal space by specialized cells, the scleroblasts, adjacent to the basal epidermal cell layer. Then, this matrix is mineralized and forms a bone devoid of cells. This type of bone is called dermal, or intramembranous, and contrarily to endochondral bone, no cartilage precursor precedes its formation. The matrix secretion and mineralization follow the proximal to distal progression of the fin growth.
Although zebrafish fin constitutes a good model for fin ray morphogenesis analysis during larval development, it has been more studied during another process, regeneration. Teleost fish, like zebrafish, possess the ability to regenerate their fins, and the ablation of any part of the fin distal to the first segment will trigger a regeneration program that will give rise to a new structure identical to the amputated one.
Regeneration in zebrafish is epimorphic, which means it involves cell proliferation and creation of a regeneration-specific structure, the blastema. In many aspects, the regeneration process is reminiscent of the development, and most of the genes expressed during embryonic or larval development are reexpressed during regeneration.
The regeneration process can be divided into three main steps32-35 (fig. 4B-D):
- Wound healing (0-24hpac): Within the 6 hours post amputation (hpa), an epithelial layer completely covers the wound, followed in the next hours by several additional layers of epidermal tissue. This forms the apical epidermal cap (AEC, fig. 4B) in a process that does not involve cell proliferation, but migration of epithelial cells from the unamputated region.36,37 The innermost cell layer (the basal epidermal layer) located against the mesenchyme, recognizable by the cuboidal shape of cells, differentiates quickly after the formation of the AEC and is the source of factors regulating epithelial-mesenchymal interactions which will control the regenerate outgrowth.15,34,38,39
- Blastema formation (24-72hpa). Following the formation of the AEC, fibroblast-like cells located up to two segments proximally to the amputation plane start to disorganize and migrate to the distal region, at the site where the blastema will form by cell proliferation.34,36,37 It is still unclear whether these cells originate from the dedifferentiation of preexisting mesenchymal cells or from a population of progenitor cells. The blastema becomes clearly visible by 2dpa. At that time, mesenchymal cell division mostly occurs in the blastema region, whereas proliferating epithelial cells are restricted to more proximal regions of the fin.37
- Blastema maturation and regenerative outgrowth (72hpa and later). Immediately after the blastema formation, mesenchymal cells segregate into three populations (fig. 4E): First, a small population of slow-cycling cells is located in the distal blastema (DB).37,40 It has been proposed that this population would constitute a pool of undifferentiated progenitor cells for the second population in the proximal blastema region (PB) which shows an intense and rapid cell cycling, twice as fast as during blastema formation.37 Finally, the most proximal part of the regenerate, the patterning zone (PZ), is mostly composed of differentiating mesenchymal cells in the core of the regenerate and scleroblasts in the periphery, adjacent to the basal epidermal layer. Cells of the PZ show little or no cell division. As regeneration continues, the blastema constantly remains distally located, driven by cell proliferation occurring in the PB, while cells of the PZ progressively differentiate into new structures which replace the amputated part of the fin (fig. 4C,D). Complete regeneration is achieved within 3 weeks depending on the amputation level.
The Hedgehog Pathway and Fin Ray Patterning: Role of the Epithelial-Mesenchymal Interactions
shh-expressing cells are detected in a broad domain covering the distal tips of each ray at about 30hpa. At 2dpa, the expression localizes to cells of the basal epidermal layer (BEL) that covers the whole surface of the amputated ray. ptc1 is also detected by 40hpa in the distal BEL, consistent with its role as a mediator of the Hh signal. During the outgrowth phase, starting by 4dpa, shh expression becomes restricted to a subset of cells of the BEL adjacent to the newly formed lepidotrichia, at the level of the PB and the PZ of the regenerate in each hemiray15,39 (fig. 4H). As regeneration proceeds, shh domain of expression regularly splits into two discrete cell populations in each hemiray (fig. 4F). This event always precedes the morphological bifurcation of the fin ray, suggesting a possible role for shh in the specification of the bifurcation. However, ptc1, which is expressed at that stage in the BEL at the level of the shh-expressing cells and also in the adjacent scleroblasts, always show a single domain of expression spanning the entire width of the hemiray (fig. 4G). This raises the possibility that factors expressed in between the two shh domains would inhibit shh signal in the central region of the hemiray in the early steps of branching formation.
A second Hh gene, coding for an orthologue of the mammalian Indian hedgehog (Ihh), was recently isolated in our laboratory.4 ihh-expressing cells are observed at 4dpa in the scleroblasts expressing ptc1 and adjacent to the shh-expressing cells of the BEL (fig. 4I). The fact that ihh is transcribed in the differentiating scleroblasts may suggest a more direct role for this factor in bone formation than shh. Furthermore, this expression of ihh in scleroblasts is unexpected, as this gene has previously been shown to be expressed in cartilage cells during endochondral bone formation only.41,42
Due to its easy access and relative simplicity, the zebrafish fin ray is a good model to perform functional and genetic studies of the regeneration process. However, as only few mutants survive long enough to be studied during fin regeneration, it has been necessary to develop and adapt new methods to manipulate gene activity in this system. Chemical treatments,15,34,43 cell transfection by microinjection43 and temperature-inducible mutants32,37,44 have revealed the requirement of the Hh signaling for proper patterning of the bony rays during fin regeneration.
RA treatments of zebrafish undergoing fin regeneration cause an inhibition of the regenerate growth followed by ray patterning defects.45 Treatments as short as 12h transiently inhibit regenerate outgrowth and downregulate shh expression. Thus, as in the embryonic fin buds,14 RA treatments of regenerating fins lead to a rapid downregulation of shh expression, supporting the idea of a direct role of RA in shh transcription via the RARE located in 5' region of the shh locus. After the end of RA treatment, the regenerate outgrowth resumes almost immediately whereas it takes 3 days for shh transcripts to be detected again. When reinitiated, the distal limit of the shh expression domain corresponds to the distal limit of the bone matrix deposition suggesting that shh expression may determine some aspects of scleroblast differentiation and patterning.15
Further evidence of the role of the Hh pathway in bone patterning has been brought by Quint et al who developed a method of gene transfection based on microinjection of plasmid DNA into the blastema (fig. 5).43 Ectopic expression of shh following injection of plasmid constructs coding for the active peptide of shh between ray branches induced an ectopic expression of ptc1 in this region and the fusions of the two branches. These fusions are caused by deposition of ectopic bone material between the basal epidermal layer and the mesenchyme of the interray region (fig. 5C-D). However, no bone forms in the deeper mesenchyme of the blastema where the shh transgene is also expressed, as it is indicated by the induction of ptc1. This suggests that only cells at the epithelial-mesenchyme interface have the potential to differentiate into scleroblasts. Bone morphogenetic proteins (BMP), members of the transforming growth factors β (TGF-β) family, are able to promote bone formation in both in vivo and in vitro systems.46,47 During fin regeneration, bmp2b is expressed in the distal BEL in a pattern similar to that of shh, as well as in the adjacent scleroblasts.15 A second member of the BMP family, bmp4, is restricted to the distal mesenchyme. bmp2b ectopic expression analysis using the approach described above leads to bone fusions similar to those obtained following ectopic shh expression (fig. 5E-F). However, cotransfection of shh and chordin, an inhibitor of the BMP signal,48 fails to produce any fusion. Altogether, these results indicate that the effect of shh ectopic expression is mediated by BMP signaling, which would act downstream of shh.43 As bmp2b ectopic expression does not induce shh-dependent ptc1 expression, no feedback loop mechanism between BMP and Hh seems to exist in the regenerate.
The effects of a loss of Hh signaling in the fin regenerate were analyzed using the steroidal alkaloid, cyclopamine, an inhibitor of Hh signaling.49 Treatments of regenerating fins with cyclopamine initially cause a proximal extension of the expression domain of shh, with a slight reduction of the regenerate outgrowth. In a second step, the outgrowth is completely inhibited and shh is no longer expressed. After 5 days of treatment, the regenerate is much shorter compared to an untreated fin, shows an accumulation of pigment cells in the distal region and no ray bifurcation. The initial upregulation of shh is suggestive of a feedback mechanism that would normally restrict shh expression at the level of the PB and PZ. The progressive arrest of fin regeneration, correlates with an inhibition of blastema cell proliferation in the regenerate epidermis and mesenchyme suggesting that the Hh pathway is necessary for blastema maintenance and outgrowth. Interestingly, bone deposition is still taking place but with abnormal patterns suggesting that inhibition of Hh signaling does not affect already-differentiated scleroblasts, but may rather affect the proliferation of undifferentiated blastema cells, their survival, and/or differentiation into specialized cell types, including scleroblasts. Another possible role for the Hh signals could be the regulation of the distribution of scleroblasts, i.e., their alignment against the basal epidermal layer. This role is further suggested by the phenotype of the temperature-sensitive regeneration mutant, emmental (emm). This mutant, in which sly1 (a gene coding for a protein involved in protein trafficking) is disrupted when fish are subjected to a heat shock at 33°C, presents blastema formation defects and a downregulation of shh expression.50 Interestingly, scleroblasts of regenerating emm fins are no longer ordered against the BEL but randomly dispersed throughout the blastema, a phenotype which connects scleroblast alignment and shh signaling.
It is likely that the fibroblast growth factors (Fgf ) play an important role in fin regeneration, possibly through interaction with the Hh pathway. Wfgf/fgf2451 a Fgf ligand of the fgf8/fgf17/fgf18 subclass, and the receptor fgfr134 are expressed during fin regeneration. fgfr1 is expressed at 18hpa in the forming blastema, then during the outgrowth phase in the distal BEL (including shh-expressing cells) and the distal blastema.34 wfgf expression is restricted to the epithelium at the distal part of the regenerate and appears relatively late, at 48hpa, suggesting that an additional, yet unidentified, Fgf ligand could be expressed at earlier stage concurrently with fgfr1. Interactions between the Fgf-Hh signaling pathways are probable, as treatments of regenerating fins with the Fgf signal inhibitor SU5402 causes effects reminiscent of cyclopamine treatments: blastema outgrowth inhibition and down-regulation of shh without affecting the wound epidermis or bone deposition.
The Wnt pathway is also involved in regeneration and is likely interacting with the Hh pathway. Wnt factors are secreted molecules, unrelated to the Hh proteins but sharing a lot of similarities with them at the structural level and their mode of action.52,53 Complex regulations exist between the Wnt and Hh pathways depending on the tissue in which they are expressed. For example, ectopic activation of β-catenin, which transduces the wnt signal to the nucleus, induces shh expression in mouse epidermis,54 whereas wnt3 is able to counteract the effect of shh overexpression in chick neural tube explants.55 The Wnt pathway is also involved in bone formation as β-catenin is required for osteoblast (bone forming cells of the endoskeleton) differentiation in the mouse embryo, possibly by acting downstream of the Hh pathway.56 Several members of the Wnt signaling pathway, β-catenin, wnt3a, wnt5 and the transcription factor lef1, are expressed during fin regeneration.39 In the early stages, β-catenin is expressed in the distal blastema whereas wnt5 and lef1 transcripts are located in proximal cells of the wound epidermis. During fin outgrowth, wnt5 is expressed in the distal BEL, and lef1 is found in most of the BEL including shh-expressing cells. Both RA treatment and inhibition of Fgf signaling using SU5402 during fin outgrowth downregulate lef1 expression, in the same way as shh expression.39 This coregulation suggests that the Hh and Wnt pathways may participate in similar processes in the basal epidermal layer during the outgrowth phase of fin regeneration.
Concluding Remarks and Future Prospects
This chapter presented the main data related to the role of the Hedgehog pathway during fin bud development and fin regeneration. The early steps of zebrafish pectoral fin development are highly reminiscent of tetrapod limb development, and shh function appears to be conserved in this process. An in-depth observation of the fin phenotype of you-type mutants is likely to provide new insights into the role and the regulation of the Hh pathway, as it will allow us to dissect the Hh pathway, and analyze the effects of its disruption at various levels. In the regenerating fin, shh plays a role in blastema maintenance and the patterning of the regenerating bony ray, probably through the correct differentiation and alignment of the scleroblasts. Future studies will further investigate the role of the Hh pathway during regeneration and its interaction with the major signaling pathways that have been described during embryonic development.
References
- 1.
- Krauss S, Concordet JP, Ingham PW. A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell. 1993;75(7):1431–1444. [PubMed: 8269519]
- 2.
- Ekker SC, Ungar AR, Greenstein P. et al. Patterning activities of vertebrate hedgehog proteins in the developing eye and brain. Curr Biol. 1995;5(8):944–955. [PubMed: 7583153]
- 3.
- Currie PD, Ingham PW. Induction of a specific muscle cell type by a hedgehog-like protein in zebrafish. Nature. 1996;382(6590):452–455. [PubMed: 8684485]
- 4.
- Avaron F, Hoffman L, Guay D. et al. Characterization of two new zebrafish members of the hedgehog family: atypical expression of the zebrafish indian hedgehog gene in skeletal elements of both endochondral and dermal origins 2005. in press . [PubMed: 16292774]
- 5.
- Dane PJ, Tucker JB. Modulation of epidermal cell shaping and extracellular matrix during caudal fin morphogenesis in the zebra fish Brachydanio rerio. J Embryol Exp Morphol. 1985;87:145–161. [PubMed: 4031750]
- 6.
- Grandel H, Schulte-Merker S. The development of the paired fins in the zebrafish (Danio rerio). Mech Dev. 1998;79(1-2):99–120. [PubMed: 10349624]
- 7.
- Akimenko MA, Ekker M. Anterior duplication of the Sonic hedgehog expression pattern in the pectoral fin buds of zebrafish treated with retinoic acid. Dev Biol. 1995;170(1):243–247. [PubMed: 7601313]
- 8.
- Riddle RD, Johnson RL, Laufer E. et al. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell. 1993;75(7):1401–1416. [PubMed: 8269518]
- 9.
- Fallon JF, Lopez A, Ros MA. et al. FGF-2: Apical ectodermal ridge growth signal for chick limb development. Science. 1994;264:104–107. [PubMed: 7908145]
- 10.
- Vogel A, Rodriguez C, Izpisúa-Belmonte J-C. Involvement of FGF-8 in initiation, outgrowth and patterning of the vertebrate limb. Development. 1996;122:1737–1750. [PubMed: 8674413]
- 11.
- Loomis CA, Harris E, Michaud J. et al. The mouse Engrailed-1 gene and ventral limb patterning. Nature. 1996;382:360–363. [PubMed: 8684466]
- 12.
- Riddle RD, Ensini M, Nelson C. et al. Induction of the LIM homeobox gene Lmx1 by WNT7a establishes dorsoventral pattern in the vertebrate limb. Cell. 1995;83:631–640. [PubMed: 7585966]
- 13.
- Bruneau S, Rosa FM. Dynamo, a new zebrafish DVR member of the TGF-beta superfamily is expressed in the posterior neural tube and is up-regulated by Sonic hedgehog. Mech Dev. 1997;61(1-2):199–212. [PubMed: 9076689]
- 14.
- Hoffman L, Miles J, Avaron F. et al. Exogenous retinoic acid induces a stage-specific, transient and progressive extension of Sonic hedgehog expression across the pectoral fin bud of zebrafish. Int J Dev Biol. 2002;46(7):949–956. [PubMed: 12455633]
- 15.
- Laforest L, Brown CW, Poleo G. et al. Involvement of the sonic hedgehog, patched 1 and bmp2 genes in patterning of the zebrafish dermal fin rays. Development. 1998;1251(21):4175–4184. [PubMed: 9753672]
- 16.
- Chandrasekhar A, Warren JrJT, Takahashi K. et al. Role of sonic hedgehog in branchiomotor neuron induction in zebrafish. Mech Dev. 1998;76(1-2):101–115. [PubMed: 9767138]
- 17.
- Du SJ, Dienhart M. Zebrafish tiggy-winkle hedgehog promoter directs notochord and floor plate green fluorescence protein expression in transgenic zebrafish embryos. Dev Dyn. 2001;222(4):655–666. [PubMed: 11748834]
- 18.
- Ng JK, Tamura K, Buscher D. et al. Molecular and cellular basis of pattern formation during vertebrate limb development. Curr Top Dev Biol. 1999;41:37–66. [PubMed: 9784972]
- 19.
- Capdevila J, Izpisua BelmonteJC. Patterning mechanisms controlling vertebrate limb development. Annu Rev Cell Dev Biol. 2001;17:87–132. [PubMed: 11687485]
- 20.
- Lu H-C, Revelli J-P, Goering L. et al. Retinoid signaling is required for the establishment of a ZPA and for the expression of Hoxb-8, a mediator of ZPA formation. Development. 1997;124:1643–1651. [PubMed: 9165113]
- 21.
- Helms J, Thaller C, Eichele G. Relationship between retinoic acid and sonic hedgehog, two polarizing signals in the chick wing bud. Development. 1994;120:3267–3274. [PubMed: 7720566]
- 22.
- Tickle C, Alberts BM, Wolpert L. et al. Local application of retinoic acid to the limb bud mimics the action of the polarizing region. Nature. 1982;296:564–565. [PubMed: 7070499]
- 23.
- Chang DT, Lopez A, von Kessler DP. et al. Products, genetic linkage, and limb patterning activity of a murine hedgehog gene. Development. 1994;120:3339–3353. [PubMed: 7720571]
- 24.
- Neumann CJ, Grandel H, Gaffield W. et al. Transient establishment of anteroposterior polarity in the zebrafish pectoral fin bud in the absence of sonic hedgehog activity. Development. 1999;126(21):4817–4826. [PubMed: 10518498]
- 25.
- Chang B-E, Blader P, Fischer N. et al. Axial (HNF3β) and retinoic acid receptors are regulators of the zebrafish sonic hedgehog promoter. EMBO J. 1997;16:3955–3964. [PMC free article: PMC1170019] [PubMed: 9233805]
- 26.
- Schauerte HE, van EedenFJ, Fricke C. et al. Sonic hedgehog is not required for the induction of medial floor plate cells in the zebrafish. Development. 1998;125(15):2983–2993. [PubMed: 9655820]
- 27.
- van EedenFJ, Granato M, Schach U. et al. Mutations affecting somite formation and patterning in the zebrafish, Danio rerio. Development. 1996;123:153–164. [PubMed: 9007237]
- 28.
- Barresi MJ, Stickney HL, Devoto SH. The zebrafish slow-muscle-omitted gene product is required for Hedgehog signal transduction and the development of slow muscle identity. Development. 2000;127(10):2189–2199. [PubMed: 10769242]
- 29.
- van EedenFJM, Granato M, Schach U. et al. Genetic analysis of fin formation in the zebrafish, Danio rerio. Development. 1996;123:255–262. [PubMed: 9007245]
- 30.
- Grandel H, Draper BW, Schulte-Merker S. Dackel acts in the ectoderm of the zebrafish pectoral fin bud to maintain AER signaling. Development. 2000;1271(19):4169–4178. [PubMed: 10976049]
- 31.
- Borday V, Thaeron C, Avaron F. et al. Evx1 transcription in bony fin rays segment boundaries leads to a reiterated pattern during zebrafish fin development and regeneration. Developmental Dynamics. 2001;220:91–98. [PubMed: 11169842]
- 32.
- Johnson SL, Weston JA. Temperature-sensitive mutations that cause stage-specific defects in zebrafish fin regeneration. Genetics. 1995;141:1583–1595. [PMC free article: PMC1206889] [PubMed: 8601496]
- 33.
- Poss KD, Keating MT, Nechiporuk A. Tales of regeneration in zebrafish. Developmental Dynamics. 2003;226(2):202–210. [PubMed: 12557199]
- 34.
- Poss KD, Shen J, Nechiporuk A. et al. Roles for Fgf signaling during zebrafish fin regeneration. Dev Biol. 2000;222(2):347–358. [PubMed: 10837124]
- 35.
- Akimenko MA, Mari-Beffa M, Becerra J. et al. Old questions, new tools, and some answers to the mystery of fin regeneration. Developmental Dynamics. 2003;226(2):190–201. [PubMed: 12557198]
- 36.
- Poleo G, Brown CW, Laforest L. et al. Cell proliferation and movement during earlin fin regeneration in zebrafish. Dev Dynamics. 2001;221:380–390. [PubMed: 11500975]
- 37.
- Nechiporuk A, Keating MT. A proliferation gradient between proximal and msxb-expressing distal blastema directs zebrafish fin regeneration. Development. 2002;129(11):2607–2617. [PubMed: 12015289]
- 38.
- Akimenko M-A, Johnson SL, Westerfield M. et al. Differential induction of four msx homeobox genes during fin development and regeneration in zebrafish. Development. 1995;121:347–357. [PubMed: 7768177]
- 39.
- Poss KD, Shen J, Keating MT. Induction of lef1 during zebrafish fin regeneration. Dev Dyn. 2000;219(2):282–286. [PubMed: 11002347]
- 40.
- Santamaría JA, Marí-Beffa M, Santos-Ruiz L. et al. Incorporation of bromodeoxyuridine in regenerating fin tissue of the goldfish Carassius auratus. The J of Exp Zool. 1996;275:300–307. [PubMed: 8759927]
- 41.
- St-Jacques B, Hammerschmidt M, McMahon AP. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999;13(16):2072–2086. [PMC free article: PMC316949] [PubMed: 10465785]
- 42.
- Vortkamp A, Lee K, Lanske B. et al. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science. 1996;273(5275):613–622. [PubMed: 8662546]
- 43.
- Quint E, Smith A, Avaron F. et al. Bone patterning is altered in the regenerating zebrafish caudal fin after ectopic expression of sonic hedgehog and bmp2b or exposure to cyclopamine. Proc Natl Acad Sci USA. 2002;99(13):8713–8718. [PMC free article: PMC124364] [PubMed: 12060710]
- 44.
- Poss KD, Nechiporuk A, Hillam AM. et al. Mps1 defines a proximal blastemal proliferative compartment essential for zebrafish fin regeneration. Development. 2002;129(22):5141–5149. [PubMed: 12399306]
- 45.
- Géraudie J, Brulfert A, Monnot M-J. et al. Teratogenic and morphogenetic effects of retinoic acid on the regenerating pectoral fin in zebrafish. J Exp Zool. 1994;269:12–22.
- 46.
- Canalis E, Economides AN, Gazzerro E. Bone morphogenetic proteins, their antagonists, and the skeleton. Endocr Rev. 2003;24(2):218–235. [PubMed: 12700180]
- 47.
- Yoon BS, Lyons KM. Multiple functions of BMPs in chondrogenesis. J Cell Biochem. 2004;93(1):93–103. [PubMed: 15352166]
- 48.
- Piccolo S, Sasai Y, Lu B. et al. Dorsoventral patterning in Xenopus: Inhibition of ventral signals by direct binding of chordin to BMP-4. Cell. 1996;86(4):589–598. [PMC free article: PMC3070603] [PubMed: 8752213]
- 49.
- Incardona JP, Gaffield W, Kapur RP. et al. The teratogenic Veratrum alkaloid cyclopamine inhibits sonic hedgehog signal transduction. Development. 1998;125(18):3553–3562. [PubMed: 9716521]
- 50.
- Nechiporuk A, Poss KD, Johnson SL. et al. Positional cloning of a temperature-sensitive mutant emmental reveals a role for sly1 during cell proliferation in zebrafish fin regeneration. Dev Biol. 2003;258(2):291–306. [PubMed: 12798289]
- 51.
- Fischer S, Draper BW, Neumann CJ. The zebrafish fgf24 mutant identifies an additional level of Fgf signaling involved in vertebrate forelimb initiation. Development. 2003;130(15):3515–3524. [PubMed: 12810598]
- 52.
- Nusse R. Wnts and Hedgehogs: Lipid-modified proteins and similarities in signaling mechanisms at the cell surface. Development. 2003;130(22):5297–5305. [PubMed: 14530294]
- 53.
- Watt FM. Unexpected Hedgehog-Wnt interactions in epithelial differentiation. Trends Mol Med. 2004;10(12):577–580. [PubMed: 15567325]
- 54.
- Callahan CA, Oro AE. Monstrous attempts at adnexogenesis: Regulating hair follicle progenitors through Sonic hedgehog signaling. Curr Opin Genet Dev. 2001;11(5):541–546. [PubMed: 11532396]
- 55.
- Robertson CP, Braun MM, Roelink H. Sonic hedgehog patterning in chick neural plate is antagonized by a Wnt3-like signal. Dev Dyn. 2004;229(3):510–519. [PubMed: 14991707]
- 56.
- Hu H, Hilton MJ, Tu X. et al. Sequential roles of Hedgehog and Wnt signaling in osteoblast development. Development. 2005;132(1):49–60. [PubMed: 15576404]
- 57.
- Karlstrom RO, Talbot WS, Schier AF. Comparative synteny cloning of zebrafish you-too: Mutations in the Hedgehog target gli2 affect ventral forebrain patterning. Genes Dev. 1999;13(4):388–393. [PMC free article: PMC316467] [PubMed: 10049354]
- 58.
- Nakano Y, Kim HR, Kawakami A. et al. Inactivation of dispatched 1 by the chameleon mutation disrupts Hedgehog signalling in the zebrafish embryo. Dev Biol. 2004;269(2):381–392. [PubMed: 15110707]
- 59.
- Baxendale S, Davison C, Muxworthy C. et al. The B-cell maturation factor Blimp-1 specifies vertebrate slow-twitch muscle fiber identity in response to Hedgehog signaling. Nat Genet. 2004;36(1):88–93. [PubMed: 14702044]
- 60.
- Varga ZM, Amores A, Lewis KE. et al. Zebrafish smoothened functions in ventral neural tube specification and axon tract formation. Development. 2001;128(18):3497–3509. [PubMed: 11566855]
- 61.
- Sekimizu K, Nishioka N, Sasaki H. et al. The zebrafish iguana locus encodes Dzip1, a novel zinc-finger protein required for proper regulation of Hedgehog signaling. Development. 2004;131(11):2521–2532. [PubMed: 15115751]
- 62.
- Karlstrom RO, Tyurina OV, Kawakami A. et al. Genetic analysis of zebrafish gli1 and gli2 reveals divergent requirements for gli genes in vertebrate development. Development. 2003;130(8):1549–1564. [PubMed: 12620981]
Footnotes
- a
in Xenopus laevis, ihh and dhh have respectively been named banded hedgehog (bhh) and cephalic hedgehog (chh).2
- b
This point is further described in another chapter of this book.
- c
hpa: hour-post-amputation and dpa: day-post-amputation at 28.5 degrees Celsius. Regeneration can be conducted at 33°C and results in speeding up the process up to two times compared to the standard temperature used to raise zebrafish.
- The Zebrafish Hedgehog Genes
- Overview of the Zebrafish Pectoral Fin Bud Development
- Shh and Twhh Expression during Fin Bud Development
- Shh, Retinoic Acid Regulation and the ZPA
- Mutants of the Hh Pathway and Fin Bud Development
- Fin Ray Morphogenesis and Regeneration
- The Hedgehog Pathway and Fin Ray Patterning: Role of the Epithelial-Mesenchymal Interactions
- Concluding Remarks and Future Prospects
- References
- Sonic Hedgehog Signaling in the Developing and Regenerating Fins of Zebrafish - ...Sonic Hedgehog Signaling in the Developing and Regenerating Fins of Zebrafish - Madame Curie Bioscience Database
- Muscle Morphogenesis: The Process of Embryonic Myoblast Fusion - Madame Curie Bi...Muscle Morphogenesis: The Process of Embryonic Myoblast Fusion - Madame Curie Bioscience Database
- Development of the Larval Somatic Musculature - Madame Curie Bioscience DatabaseDevelopment of the Larval Somatic Musculature - Madame Curie Bioscience Database
- Tissue Engineering Constructs and Commercialization - Madame Curie Bioscience Da...Tissue Engineering Constructs and Commercialization - Madame Curie Bioscience Database
- Electrophysiological Simulation of Developmental Changes in Action Potentials of...Electrophysiological Simulation of Developmental Changes in Action Potentials of Cardiomyocytes - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...