

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Eukaryotic gene expression involves a number of interlinked post-transcriptional steps that are subject to surveillance or quality control mechanisms to ensure that only fully processed and error-free mRNAs are translated. Among these, the nonsense-mediated mRNA decay (NMD) pathway recognizes and targets for degradation mRNAs containing premature translation termination codons (PTCs), which could give rise to truncated and potentially harmful proteins. The NMD pathway not only prevents the accumulation of malfunctioning proteins but also modulates the clinical manifestations of many human genetic disorders (see chapters by Holbrook et al and Keeling et al). A cross-species analysis of this pathway has revealed important conserved key components, and has provided the basis for elucidating the NMD network in humans. This chapter focuses on the NMD pathway in the fruitfly Drosophila melanogaster, emphasizing how studies in this model organism have provided new insights into the mechanisms underlying NMD and its evolution.
Introduction
The gene expression pathway involves a number of interlinked steps that begin with gene transcription and end with protein synthesis, wherein mRNA is a key intermediate. mRNA is transcribed from DNA as pre-mRNA, which is generally processed by the removal of introns and the addition of the poly(A) tail. The resulting mature mRNA is exported to the cytoplasm where it is translated and finally degraded. It is now established that modulation of most, if not all, steps in the gene expression pathway can contribute to the regulation of gene expression.
While the complexity of eukaryotic gene expression allows for the control of protein production at multiple levels, it also makes the process vulnerable to errors. Eukaryotic cells have evolved elaborated mRNA quality control (also called surveillance) mechanisms that ensure the fidelity of gene expression by detecting and degrading aberrant transcripts. These quality control or surveillance mechanisms operate in both the nucleus and the cytoplasm. For instance, mechanisms in the nucleus degrade improperly processed mRNAs before they are exported to the cytoplasm (reviewed by Fasken and Corbett in ref. 1). In the cytoplasm, surveillance pathways assess the translatability of the mRNA and degrade any that have no translation termination codons (nonstop-mediated mRNA decay, NSD) or that have PTCs (NMD) (reviewed by Fasken and Corbett in ref. 1).
The NMD pathway is one of the best characterized mRNA surveillance mechanisms. It not only eliminates mRNAs containing frameshift or nonsense mutations, but also regulates the expression of naturally occurring transcripts having features recognized by the NMD machinery (see chapters by He and Jacobson, Sharifi and Dietz, and Soergel et al). In this way, the NMD pathway contributes to the post-transcriptional regulation of about 10% of the transcriptome in yeast, fruitfly and human cells.2-5
Studies using model organisms have revealed that despite conservation of this pathway, different species have evolved different mechanisms to discriminate natural from premature translation termination codons and to degrade transcripts identified as NMD substrates (reviewed by Conti and Izaurralde in ref. 6). In this chapter, we discuss the information gathered from studies of the NMD pathway in the model organism Drosophila melanogaster (fruitfly) and the insights gained from these studies into the molecular mechanism underlying NMD and its evolution.
The NMD Protein Interaction Network: An Overview
The RNA helicase Upf1 is a key molecular component of the NMD machinery. Upf1 was originally identified in genetic screens of yeast (Saccharomyces cerevisiae) and worms (Caenorhabditis elegans) and shown to be essential for NMD in these organisms (see chapter by Baker and Parker and Anderson). Subsequently, it was shown that silencing of the Upf1 gene by RNA interference (RNAi) in both human and fruitfly cells results in the stabilization of PTC-containing mRNAs indicating that these Upf1 orthologs are also essential for NMD.7,8 Upf1 interacts with two additional proteins, Upf2 and Upf3, to form the so-called ‘surveillance complex’. Surveillance complex function in NMD is conserved in all organisms that have been studied. Indeed data suggests that for all eukaryotes recognition of a PTC by a translating ribosome leads to the assembly of the surveillance complex, which in turn targets the mRNA for decay.
The role of the surveillance complex is therefore to couple a premature translation termination event to mRNA degradation. Consistent with this, NMD factors have genetic, functional and/or physical links with translation factors and mRNA decay enzymes (Fig. 1). A link between NMD and translation termination is provided by the association of Upf1, Upf2 and Upf3 with the eukaryotic translation termination factors (eRF)1 and eRF3.9-11 As the actual substrate of NMD is not a naked mRNA, but a ribonucleoprotein particle (mRNP), many RNA binding proteins play roles in NMD. For yeast, these proteins include the poly(A)-binding protein 1 (Pab1p) (see chapter by Amrani and Jacobson). For mammals, they include the exon junction complex (EJC), which is a multi-protein assembly deposited ˜20-24 nucleotides upstream of exon-exon junctions (see chapter by Maquat).
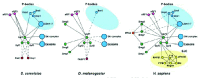
As mentioned above, although the core of the NMD machinery (Upf1-3) is conserved, the mechanisms by which PTCs are recognized and mRNA targets are degraded differ between species (reviewed by Conti and Izaurralde in ref. 6). These mechanistic differences correlate with changes in the NMD protein interaction network (Fig. 1). Moreover, the complexity of the network underlying this pathway increases from simpler organisms such as the budding yeast S. cerevisiae to the mammal Homo sapiens. For instance, additional factors have been integrated into the network in multicellular organisms, including the proteins Smg1, Smg5, Smg6 and Smg7, which regulate Upf1 function.
Upf1 is regulated by phosphorylation and dephosphorylation. In man and worms, phosphorylation of Upf1 is catalyzed by Smg1, a phosphoinositide-3-kinase-related protein kinase, and requires Upf2 and Upf312-17 (see chapters by Yamashita et al and Anderson). This suggests that it is the assembly of the surveillance complex that triggers Upf1-phosphorylation. The dephosphorylation of Upf1 is mediated by Smg5, Smg6 and Smg7, three similar but not functionally redundant proteins.14,18-20 Smg5-7 are not phosphatases themselves, but they are thought to trigger Upf1 dephosphorylation by recruiting protein phosphatase 2A (PP2A). This model is based on the observations that Smg5 and Smg7 interact with each other and are part of a larger complex comprising phosphatase PP2A and phosphorylated Upf1.19,20 Similarly, Smg6 is part of a protein complex comprising PP2A and phosphorylated Upf1.21 With the exception of Smg7, Drosophila orthologs to core components of the NMD machinery have been identified, including Upf1, Upf2, Upf3, Smg1, Smg5 and Smg6.7 Depletion of these proteins by RNAi in Drosophila Schneider cells stabilizes various PTC-containing mRNAs, providing evidence for a role in NMD.7 Intriguingly, mutations in the smg1 gene do not appear to affect NMD in Drosophila embryos.22 These observations suggest that different and/or redundant kinases phosphorylate Upf1 in embryos.
Regardless of whether Upf1 is phosphorylated by another redundant kinase in embryos, the role of Drosophila Smg1 in NMD is well established. Firstly, depletion of this protein stabilizes different PTC-containing reporters in Schneider cells to a similar extent as does depletion of Upf3.7 Secondly, depletion of Smg1 or depletion of any of the other known NMD factors leads to similar mRNA expression profiles.5 Thus, in addition of Upf1-3, NMD in Drosophila requires the Smg1, Smg5 and Smg6 proteins, which are a specific feature of higher eukaryotic NMD.
Mechanism of PTC Definition in Drosophila
In mammals, PTC recognition relies on splicing. In this case, stop codons are defined as premature if they are located ˜50 nucleotides or more upstream of an exon-exon junction23 (see chapter by Maquat). The positions of exon-exon junctions are communicated to translating ribosomes by the EJC (see chapter by Maquat) (Fig. 2). In particular, the EJC components Y14, Magoh, eIF4AIII, Barentsz and RNPS1 elicit NMD when bound at least ˜20-25 nucleotides downstream of a termination codon24-27 (Fig. 2).

Mammalian Upf3 interacts with components of the EJC and is loaded onto mRNAs during splicing, while Upf2 is thought to join the complex in the cytoplasm after export.26,28-31 During a pioneer round of translation, Upf2, Upf3 and the additional EJC components are displaced by the ribosomes as they traverse the mRNA.32 If translating ribosomes were to encounter a stop codon upstream of an EJC, this would lead to the incomplete removal of Upf2 and Upf3 proteins from downstream mRNA sequences and to the recruitment of Upf1, probably via interactions with the eRF1:eRF3 complex. The recruitment of Upf1 creates an opportunity for the assembly of the Upf1:Upf2:Upf3 complex, which targets the mRNA for rapid degradation (reviewed by Wagner and Lykke-Andersen,33 see chapters by Maquat, and Singh and Lykke-Andersen).
In S. cerevisiae, most genes lack introns, and most EJC components do not exist. Consistent with this, exon-exon junctions are not required to distinguish natural from premature stop codons in this organism. Two models have been proposed to explain the mechanism of PTC recognition in yeast. In one model, PTCs are defined relative to a downstream sequence element (DSE), which might function analogously to a mammalian exon-exon junction (see chapters by Baker and Parker, and Amrani and Jacobson). The protein Hrp1p has been implicated in binding to at least one DSE, thereby providing positional information for PTC recognition.
An alternative, the faux 3'-untranslated region (UTR) model, posits that the process of premature translation termination is intrinsically aberrant, because the stop codon is not in the appropriate context (i.e., not flanked by a bona fide 3'-UTR [see chapter by Amrani and Jacobson]). According to this model, bona fide 3'-UTRs would be marked by a specific protein or set of proteins (e.g., Pab1p). If a terminating ribosome is able to interact with these proteins, proper termination can occur. In contrast, translation termination at a PTC would be impaired or too slow, because of the inability of the terminating ribosome to establish interactions with bona fide 3'-UTR-associated proteins. In this case the NMD complex could be assembled, leading to rapid degradation of the mRNA.
Several lines of evidence support the faux 3'-UTR model in yeast. Firstly, translation termination is indeed aberrant at PTCs compared to normal termination codons, and prematurely terminating ribosomes fail to release efficiently.34 Secondly, this effect is abolished when the PTC is flanked by a normal 3'-UTR.34 Moreover, tethering Pab1p downstream of the PTC apparently mimics a normal 3'-UTR since it leads to efficient translation termination at the PTC and suppresses NMD.34
In Drosophila, as in yeast, PTC recognition occurs independently of exon-exon junctions, and although Drosophila orthologs of vertebrate EJC proteins do exist (i.e., Y14, Mago, eIF4AIII, Barentsz and RNPS1), they are dispensable for NMD.7 This concurs with the observation that PTC-containing mRNAs that are transcribed from intronless genes are subject to NMD in Drosophila,7 in contrast to the situation in mammals (see chapter by Maquat).
What differentiates a PTC from a normal termination codon in Drosophila? One possibility is that PTCs are defined relative to a DSE, as may be the case for some transcripts in yeast. However, the observation that transcripts derived from heterologous genes such as bacterial chloramphenicol-acetyl transferase (CAT) or green fluorescent protein (GFP) can be recognized by the NMD machinery when they carry a PTC makes unlikely the possibility that specific sequence elements mark Drosophila transcripts. Analogously to the results reported for yeast, we have recently found that tethering PABPC1 downstream of a PTC abolishes NMD in Drosophila cells (I. Behm-Ansmant and E. Izaurralde, unpublished). These findings strongly suggest that the faux 3'-UTR model provides a mechanism for PTC recognition in Drosophila, too.
The changes in the mechanisms of PTC definition described above are reflected in the NMD protein interaction network (Fig. 1). For instance, the proteins Y14, Magoh, eIF4AIII and Barentsz are part of a protein interaction module conserved in multicellular organisms, but not in budding yeast. In Drosophila, this module plays a role in mRNA localization, while in mammals, it interacts with the NMD machinery, reflecting the requirement of exon-exon junctions for PTC-recognition (see chapter by Maquat). In yeast and Drosophila, in contrast, poly(A)-binding proteins (Pab1p and its ortholog PABPC1, respectively) play a role in PTC definition and are therefore functionally linked to the NMD network in these organisms (Fig. 1).
Decay of NMD Targets in Drosophila
In eukaryotic cells, mRNAs are generally degraded via alternative pathways, each of which is initiated by removal of the poly(A)-tail by deadenylases (reviewed by Parker and Song in ref. 35). Following this first rate-limiting step, the cap structure is removed by the decapping enzymes Dcp1/Dcp2 and decapping coactivators such as the LSm1-7 complex, which makes the mRNA susceptible to digestion by the major cytoplasmic 5'-to-3' exonuclease Xrn1. Alternatively, following deadenylation, mRNAs can be degraded from their 3'-ends. Decay through the 3'-to-5' pathway requires the exosome, a multimeric complex of 3'-to-5' exonucleases, and the Ski complex, a trimeric protein complex that regulates exosome activity (reviewed by Parker and Song in ref. 35).
The enzymes involved in general mRNA decay also function in NMD. In yeast, the major decay pathway for NMD substrates involves deadenylation-independent decapping and 5'-to-3' degradation by Xrn1p36-38 (Fig. 3). This implies that one function of the surveillance complex is to bypass deadenylation, the rate-limiting step in mRNA decay, and to promote decapping directly. An alternative pathway, which also contributes to the decay of PTC-containing mRNAs, relies on accelerated deadenylation and 3'-to-5' degradation by the exosome and the Ski complex37,38 (Fig. 3). The mechanism by which NMD substrates are degraded in mammals has not been investigated in as much detail, but has been shown to occur by a mechanism similar to that described in yeast.39-41
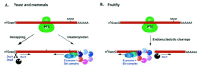
In Drosophila, in contrast, degradation of nonsense transcripts is initiated by endonucleolytic cleavage near the PTC (Fig. 3). The resulting 5' fragment is degraded by the exosome, while the 3' fragment is degraded by Xrn1.42 Thus, the mRNA fragments are degraded from the newly generated ends without undergoing decapping or deadenylation, suggesting that, in contrast to yeast and mammals, the decapping enzymes, the LSm1-7 complex and deadenylases are not required for NMD in Drosophila, and are not part of the NMD protein interaction network (Fig. 1).
Recently, the decapping enzymes Dcp1 and Dcp2, the LSm1-7 complex and Xrn1 have been shown to reside in specialized cytoplasmic bodies or mRNA decay foci.43-47 These foci have been named mRNA processing bodies (P-bodies) or GW bodies, due to the accumulation of the RNA binding protein GW182 in these bodies.44,48 These findings have raised the question of whether decay of NMD substrates occurs in the cytoplasm independently of P-bodies or whether the entire surveillance complex escorts the mRNA to P-bodies, where it undergoes rapid 5'-3' decay (see chapters by Baker and Parker, and Singh and Lykke-Andersen).
Smg7 provides a molecular link between NMD and decay enzymes localized in P-bodies.49,50 When over-expressed, human Smg7 accumulates in P-bodies and also causes the accumulation of Smg5 and Upf1 in these bodies.49,50 These observations suggest a mechanism that couples the assembly of the surveillance complex to the degradation of PTC-containing transcripts. Accordingly, the assembly of the surveillance complex on PTC-containing mRNAs leads to the phosphorylation of Upf1. Phosphorylated Upf1 recruits Smg7 (most likely in a complex with Smg5 and PP2A).19,20,49 Smg7 would then recruit the decay enzymes in the cytoplasm or target the PTC-containing transcript for degradation in P-bodies.49 The association of Smg7, Smg5 and PP2A would also trigger the dephosphorylation of Upf1, which might be involved in recycling of the NMD factors for another round of targeting.
The mechanism for connecting the surveillance complex to the mRNA decay enzymes in yeast and Drosophila cells (which appear to lack a Smg7 ortholog) has not been fully elucidated. Moreover, the identity of the Drosophila endonuclease and how it is recruited to nonsense transcripts are unknown.
Evolution of the Physiological Role of NMD
NMD components are not essential in yeast.51 Similarly, inhibition of the NMD pathway in C. elegans leads to viable worms with defects in the male bursa and the hermaphrodite vulva16,52 (see chapter by Anderson). In contrast, depletion of Upf1 or Upf2 induces a G2/M cell cycle arrest and impairs cell proliferation in Drosophila.5 Upf1 is also required for cell viability in mice, and deletion of the UPF1 gene leads to early embryonic death.53
At least two mechanisms can account for the different phenotypes observed across species after depletion of NMD factors. One mechanism could be the acquisition of novel functions by NMD components during evolution. Consistent with this possibility, it has been reported that human Upf1 can be recruited to the 3' UTR of specific transcripts via interactions with Staufen1 or stem-loop binding protein and elicits mRNA decay by a mechanism distinct from but related to NMD54 (see chapters by Kim and Maquat, and Kaygun and Marzluff). Moreover, recent reports have implicated a subset of NMD factors in other cellular processes including telomere maintenance and DNA repair (see chapters by Abraham and Oliveira, and Azzalin et al).
Another mechanism that can lead to phenotypic differences is changes in selected targets. Evidence already exists that this is indeed the case. For instance, rearrangements of the immunoglobulin and T-cell receptor genes in vertebrates result in frame-shifted genes at high frequency (˜66% of the recombination events), and transcripts from these genes are degraded by NMD (see chapter by Gudikote and Wilkinson). More recently gene expression profiling of yeast, fruitfly or human cells defective in NMD have allowed the identification of endogenous targets of the pathway (see ref. 5 and chapters by He and Jacobson, Sharafi and Dietz, and Soergel et al). These studies revealed that although NMD regulates the expression of ˜10% of the transcriptome in these organism,2-5 with few exceptions, NMD targets in different species do not represent orthologous genes.5
The exceptions include transcripts involved in DNA repair and telomere maintenance. SMG5 mRNA is regulated by NMD in both human and Drosophila cells, and in human cells it has been implicated in telomerase function.4,5,55,56 Similarly, the telomerase-associated protein Est1p (a close relative of Smg5-7) is regulated by NMD in yeast.3 In addition to EST1, NMD regulates six genes involved in telomere maintenance in yeast, including ESB1, EST2, EST3, STN1, YKu70 and TEL1.3,57 This regulation has functional implications, as inhibition of NMD in yeast leads to telomere shortening and de-repression of silenced telomeric loci.3,57,58 Finally, the gene for Drosophila ATM, a relative of TEL1 involved in DNA repair and telomere function, is also a target of NMD.5 All together, this suggests a conserved role for NMD in regulating the expression of genes involved in telomere maintenance.
The analysis of gene expression profiles in cells lacking individual NMD factors also revealed that in both yeast and fruitfly cells the major role of these factors is to regulate in concert the expression of a common set of genes. Indeed, yeast strains lacking Upf1p, Upf2p or Upf3p exhibit similar expression profiles.3 Moreover, depletion of Upf1, Upf2, Upf3, Smg1, Smg5 or Smg6 from fruitfly cells, results in correlated changes in gene expression.5 This and the observation that only a few transcripts are regulated exclusively by the individual proteins suggest that most NMD factors have not acquired specialized functions in RNA turnover in these organisms. These findings do not rule out the possibility that individual NMD factors have acquired additional functions that do not affect steady-state mRNA levels. Consequently, differences in the cellular role of NMD between yeast and fruitfly largely reflect changes in selected targets in addition to a potential functional diversification of NMD components. An important goal for the future is to understand how changes in regulated targets lead to the different phenotypes observed at the cellular level across species.
Perspectives
NMD is linked to other post-transcriptional processes by the use of common factors. For instance, many RNA binding proteins play roles at multiple steps of gene expression. As one example, poly(A)-binding proteins have roles in translation, NMD and mRNA turnover. In addition, enzymes involved in general mRNA degradation are recruited by the NMD and RNA silencing machineries to degrade targeted mRNAs. Understanding how the interplay of these regulatory circuits determines the expression pattern of many genes is an important goal for the future.
The NMD pathway contributes to the phenotypic manifestations of human genetic diseases, and therapeutic strategies based on the modulation of this pathway are under evaluation (see chapters by Holbrook et al and Keeling et al). However, the effect of such strategies on the whole patient cannot be assessed in the absence of information on the physiological role of this pathway. A deeper understanding of the mechanisms underlying NMD and the comprehensive identification of its components are also required. Studies in model organisms such as yeast, worms and fruitfly have begun to provide a detailed understanding of the mechanisms of NMD, and they have led to the identification of the core components of the machinery. These studies are likely to continue to unveil deep insights into the role and regulatory mechanisms of NMD, and they will contribute to establishing the NMD network in humans.
Acknowledgements
We gratefully acknowledge David Thomas for critical reading of the manuscript. Both authors are supported by the European Molecular Biology Organization (EMBO) and the Human Frontier Science Program Organization (HFSPO).
References
- 1.
- Fasken MB, Corbett AH. Process or perish: Quality control in mRNA biogenesis. Nat Struct Mol Biol. 2005;12(6):482–488. [PubMed: 15933735]
- 2.
- Lelivelt MJ, Culbertson MR. Yeast Upf proteins required for RNA surveillance affect global expression of the yeast transcriptome. Mol Cell Biol. 1999;19(10):6710–6719. [PMC free article: PMC84660] [PubMed: 10490610]
- 3.
- He F, Li X, Spatrick P. et al. Genome-wide analysis of mRNAs regulated by the nonsense-mediated and 5' to 3' mRNA decay pathways in yeast. Mol Cell. 2003;12(6):1439–1452. [PubMed: 14690598]
- 4.
- Mendell JT, Sharifi NA, Meyers JL. et al. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat Genet. 2004;36(10):1073–1078. [PubMed: 15448691]
- 5.
- Rehwinkel J, Letunic I, Raes J. et al. Nonsense-mediated mRNA decay factors act in concert to regulate common mRNA targets. RNA. 2005;11(10):1530–1544. [PMC free article: PMC1370837] [PubMed: 16199763]
- 6.
- Conti E, Izaurralde E. Nonsense-mediated mRNA decay: Molecular insights and mechanistic variations across species. Curr Opin Cell Biol. 2005;17(3):316–325. [PubMed: 15901503]
- 7.
- Gatfield D, Unterholzner L, Ciccarelli FD. et al. Nonsense-mediated mRNA decay in Drosophila: At the intersection of the yeast and mammalian pathways. EMBO J. 2003;22(15):3960–3970. [PMC free article: PMC169044] [PubMed: 12881430]
- 8.
- Mendell JT, ap Rhys CM, Dietz HC. Separable roles for rent1/hUpf1 in altered splicing and decay of nonsense transcripts. Science. 2002;298(5592):419–422. [PubMed: 12228722]
- 9.
- Weng Y, Czaplinski K, Peltz SW. Identification and characterization of mutations in the UPF1 gene that affect nonsense suppression and the formation of the Upf protein complex but not mRNA turnover. Mol Cell Biol. 1996;16(10):5491–5506. [PMC free article: PMC231549] [PubMed: 8816462]
- 10.
- Wang W, Czaplinski K, Rao Y. et al. The role of Upf proteins in modulating the translation read-through of nonsense-containing transcripts. EMBO J. 2001;20(4):880–890. [PMC free article: PMC145432] [PubMed: 11179232]
- 11.
- Czaplinski K, Ruiz-Echevarria MJ, Paushkin SV. et al. The surveillance complex interacts with the translation release factors to enhance termination and degrade aberrant mRNAs. Genes Dev. 1998;12(11):1665–1677. [PMC free article: PMC316864] [PubMed: 9620853]
- 12.
- Denning G, Jamieson L, Maquat LE. et al. Cloning of a novel phosphatidylinositol kinase-related kinase: Characterization of the human Smg-1 RNA surveillance protein. J Biol Chem. 2001;276(25):22709–22714. [PubMed: 11331269]
- 13.
- Grimson A, O'Connor S, Newman CL. et al. Smg-1 is a phosphatidylinositol kinase-related protein kinase required for nonsense-mediated mRNA Decay in Caenorhabditis elegans. Mol Cell Biol. 2004;24(17):7483–7490. [PMC free article: PMC506987] [PubMed: 15314158]
- 14.
- Page MF, Carr B, Anders KR. et al. Smg-2 is a phosphorylated protein required for mRNA surveillance in Caenorhabditis elegans and related to Upf1p of yeast. Mol Cell Biol. 1999;19(9):5943–5951. [PMC free article: PMC84455] [PubMed: 10454541]
- 15.
- Pal M, Ishigaki Y, Nagy E. et al. Evidence that phosphorylation of human Upfl protein varies with intracellular location and is mediated by a wortmannin-sensitive and rapamycin-sensitive PI 3-kinase-related kinase signaling pathway. RNA. 2001;7(1):5–15. [PMC free article: PMC1370068] [PubMed: 11214180]
- 16.
- Pulak R, Anderson P. mRNA surveillance by the Caenorhabditis elegans smg genes. Genes Dev. 1993;7(10):1885–1897. [PubMed: 8104846]
- 17.
- Yamashita A, Ohnishi T, Kashima I. et al. Human Smg-1, a novel phosphatidylinositol 3-kinase-related protein kinase, associates with components of the mRNA surveillance complex and is involved in the regulation of nonsense-mediated mRNA decay. Genes Dev. 2001;15(17):2215–2228. [PMC free article: PMC312771] [PubMed: 11544179]
- 18.
- Cali BM, Kuchma SL, Latham J. et al. smg-7 is required for mRNA surveillance in Caenorhabditis elegans. Genetics. 1999;151(2):605–616. [PMC free article: PMC1460488] [PubMed: 9927455]
- 19.
- Anders KR, Grimson A, Anderson P. Smg-5, required for C.elegans nonsense-mediated mRNA decay, associates with Smg-2 and protein phosphatase 2A. EMBO J. 2003;22(3):641–650. [PMC free article: PMC140740] [PubMed: 12554664]
- 20.
- Ohnishi T, Yamashita A, Kashima I. et al. Phosphorylation of hUPF1 induces formation of mRNA surveillance complexes containing hSMG-5 and hSMG-7. Mol Cell. 2003;12(5):1187–1200. [PubMed: 14636577]
- 21.
- Chiu SY, Serin G, Ohara O. et al. Characterization of human Smg5/7a: A protein with similarities to Caenorhabditis elegans SMG5 and SMG7 that functions in the dephosphorylation of Upf1. RNA. 2003;9(1):77–87. [PMC free article: PMC1370372] [PubMed: 12554878]
- 22.
- Chen Z, Smith KR, Batterham P. et al. Smg1 nonsense mutations do not abolish nonsense-mediated mRNA decay in Drosophila melanogaster. Genetics. 2005 [PMC free article: PMC1456532] [PubMed: 15965240]
- 23.
- Nagy E, Maquat LE. A rule for termination-codon position within intron-containing genes: When nonsense affects RNA abundance. Trends Biochem Sci. 1998;23(6):198–199. [PubMed: 9644970]
- 24.
- Fribourg S, Gatfield D, Izaurralde E. et al. A novel mode of RBD-protein recognition in the Y14-Mago complex. Nat Struct Biol. 2003;10(6):433–439. [PubMed: 12730685]
- 25.
- Gehring NH, Neu-Yilik G, Schell T. et al. Y14 and hUpf3b form an NMD-activating complex. Mol Cell. 2003;11(4):939–949. [PubMed: 12718880]
- 26.
- Lykke-Andersen J, Shu MD, Steitz JA. Communication of the position of exon-exon junctions to the mRNA surveillance machinery by the protein RNPS1. Science. 2001;293(5536):1836–1839. [PubMed: 11546874]
- 27.
- Palacios IM, Gatfield D, St Johnston D. et al. An eIF4AIII-containing complex required for mRNA localization and nonsense-mediated mRNA decay. Nature. 2004;427(6976):753–757. [PubMed: 14973490]
- 28.
- Kim VN, Kataoka N, Dreyfuss G. Role of the nonsense-mediated decay factor hUpf3 in the splicing-dependent exon-exon junction complex. Science. 2001;293(5536):1832–1836. [PubMed: 11546873]
- 29.
- Le Hir H, Gatfield D, Izaurralde E. et al. The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay. EMBO J. 2001;20(17):4987–4997. [PMC free article: PMC125616] [PubMed: 11532962]
- 30.
- Lykke-Andersen J, Shu MD, Steitz JA. Human Upf proteins target an mRNA for nonsense-mediated decay when bound downstream of a termination codon. Cell. 2000;103(7):1121–1131. [PubMed: 11163187]
- 31.
- Serin G, Gersappe A, Black JD. et al. Identification and characterization of human orthologues to Saccharomyces cerevisiae Upf2 protein and Upf3 protein (Caenorhabditis elegans SMG-4). Mol Cell Biol. 2001;21(1):209–223. [PMC free article: PMC88795] [PubMed: 11113196]
- 32.
- Ishigaki Y, Li X, Serin G. et al. Evidence for a pioneer round of mRNA translation: mRNAs subject to nonsense-mediated decay in mammalian cells are bound by CBP80 and CBP20. Cell. 2001;106(5):607–617. [PubMed: 11551508]
- 33.
- Wagner E, Lykke-Andersen J. mRNA surveillance: The perfect persist. J Cell Sci. 2002;115(Pt 15):3033–3038. [PubMed: 12118059]
- 34.
- Amrani N, Ganesan R, Kervestin S. et al. A faux 3'-UTR promotes aberrant termination and triggers nonsense-mediated mRNA decay. Nature. 2004;432(7013):112–118. [PubMed: 15525991]
- 35.
- Parker R, Song H. The enzymes and control of eukaryotic mRNA turnover. Nat Struct Mol Biol. 2004;11(2):121–127. [PubMed: 14749774]
- 36.
- Muhlrad D, Parker R. Aberrant mRNAs with extended 3' UTRs are substrates for rapid degradation by mRNA surveillance. RNA. 1999;5(10):1299–1307. [PMC free article: PMC1369852] [PubMed: 10573121]
- 37.
- Cao D, Parker R. Computational modeling and experimental analysis of nonsense-mediated decay in yeast. Cell. 2003;113(4):533–545. [PubMed: 12757713]
- 38.
- Mitchell P, Tollervey D. An NMD pathway in yeast involving accelerated deadenylation and exosome-mediated 3'→5' degradation. Mol Cell. 2003;11(5):1405–1413. [PubMed: 12769863]
- 39.
- Chen CY, Shyu AB. Rapid deadenylation triggered by a nonsense codon precedes decay of the RNA body in a mammalian cytoplasmic nonsense-mediated decay pathway. Mol Cell Biol. 2003;23(14):4805–4813. [PMC free article: PMC162215] [PubMed: 12832468]
- 40.
- Couttet P, Grange T. Premature termination codons enhance mRNA decapping in human cells. Nucleic Acids Res. 2004;32(2):488–494. [PMC free article: PMC373342] [PubMed: 14742663]
- 41.
- Lejeune F, Li X, Maquat LE. Nonsense-mediated mRNA decay in mammalian cells involves decapping, deadenylating, and exonucleolytic activities. Mol Cell. 2003;12(3):675–687. [PubMed: 14527413]
- 42.
- Gatfield D, Izaurralde E. Nonsense-mediated messenger RNA decay is initiated by endonucleolytic cleavage in Drosophila. Nature. 2004;429(6991):575–578. [PubMed: 15175755]
- 43.
- Cougot N, Babajko S, Seraphin B. Cytoplasmic foci are sites of mRNA decay in human cells. J Cell Biol. 2004;165(1):31–40. [PMC free article: PMC2172085] [PubMed: 15067023]
- 44.
- Eystathioy T, Jakymiw A, Chan EK. et al. The GW182 protein colocalizes with mRNA degradation associated proteins hDcp1 and hLSm4 in cytoplasmic GW bodies. RNA. 2003;9(10):1171–1173. [PMC free article: PMC1370480] [PubMed: 13130130]
- 45.
- Ingelfinger D, Arndt-Jovin DJ, Luhrmann R. et al. The human LSm1-7 proteins colocalize with the mRNA-degrading enzymes Dcp1/2 and Xrnl in distinct cytoplasmic foci. RNA. 2002;8(12):1489–1501. [PMC free article: PMC1370355] [PubMed: 12515382]
- 46.
- Sheth U, Parker R. Decapping and decay of messenger RNA occur in cytoplasmic processing bodies. Science. 2003;300(5620):805–808. [PMC free article: PMC1876714] [PubMed: 12730603]
- 47.
- van Dijk E, Cougot N, Meyer S. et al. Human Dcp2: A catalytically active mRNA decapping enzyme located in specific cytoplasmic structures. EMBO J. 2002;21(24):6915–6924. [PMC free article: PMC139098] [PubMed: 12486012]
- 48.
- Eystathioy T, Chan EK, Tenenbaum SA. et al. A phosphorylated cytoplasmic autoantigen, GW182, associates with a unique population of human mRNAs within novel cytoplasmic speckles. Mol Biol Cell. 2002;13(4):1338–1351. [PMC free article: PMC102273] [PubMed: 11950943]
- 49.
- Unterholzner L, Izaurralde E. SMG7 acts as a molecular link between mRNA surveillance and mRNA decay. Mol Cell. 2004;16(4):587–596. [PubMed: 15546618]
- 50.
- Fukuhara N, Ebert J, Unterholzner L. et al. SMG7 is a 14-3-3-like adaptor in the nonsense-mediated mRNA decay pathway. Mol Cell. 2005;17(4):537–547. [PubMed: 15721257]
- 51.
- Leeds P, Peltz SW, Jacobson A. et al. The product of the yeast UPF1 gene is required for rapid turnover of mRNAs containing a premature translational termination codon. Genes Dev. 1991;5(12A):2303–2314. [PubMed: 1748286]
- 52.
- Hodgkin J, Papp A, Pulak R. et al. A new kind of informational suppression in the nematode Caenorhabditis elegans. Genetics. 1989;123(2):301–313. [PMC free article: PMC1203802] [PubMed: 2583479]
- 53.
- Medghalchi SM, Frischmeyer PA, Mendell JT. et al. Rent1, a trans-effector of nonsense-mediated mRNA decay, is essential for mammalian embryonic viability. Hum Mol Genet. 2001;10(2):99–105. [PubMed: 11152657]
- 54.
- Kim YK, Furic L, Desgroseillers L. et al. Mammalian Staufen1 recruits Upf1 to specific mRNA 3'UTRs so as to elicit mRNA decay. Cell. 2005;120(2):195–208. [PubMed: 15680326]
- 55.
- Reichenbach P, Hoss M, Azzalin CM. et al. A human homolog of yeast Est1 associates with telomerase and uncaps chromosome ends when overexpressed. Curr Biol. 2003;13(7):568–574. [PubMed: 12676087]
- 56.
- Snow BE, Erdmann N, Cruickshank J. et al. Functional conservation of the telomerase protein Est1p in humans. Curr Biol. 2003;13(8):698–704. [PubMed: 12699629]
- 57.
- Dahlseid JN, Lew-Smith J, Lelivelt MJ. et al. mRNAs encoding telomerase components and regulators are controlled by UPF genes in Saccharomyces cerevisiae. Eukaryot Cell. 2003;2(1):134–142. [PMC free article: PMC141172] [PubMed: 12582130]
- 58.
- Lew JE, Enomoto S, Berman J. Telomere length regulation and telomeric chromatin require the nonsense-mediated mRNA decay pathway. Mol Cell Biol. 1998;18(10):6121–6130. [PMC free article: PMC109198] [PubMed: 9742129]
- NMD in Drosophila: A Snapshot into the Evolution of a Conserved mRNA Surveillanc...NMD in Drosophila: A Snapshot into the Evolution of a Conserved mRNA Surveillance Pathway - Madame Curie Bioscience Database
- A Prelude to Translational (Re)Initiation - Madame Curie Bioscience DatabaseA Prelude to Translational (Re)Initiation - Madame Curie Bioscience Database
- Adding Complexity to Phagocytic Signaling: Phagocytosis-Associated Cell Response...Adding Complexity to Phagocytic Signaling: Phagocytosis-Associated Cell Responses and Phagocytic Efficiency - Madame Curie Bioscience Database
- Asparaginyl-tRNA Synthetases - Madame Curie Bioscience DatabaseAsparaginyl-tRNA Synthetases - Madame Curie Bioscience Database
- Therapies of Nonsense-Associated Diseases - Madame Curie Bioscience DatabaseTherapies of Nonsense-Associated Diseases - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...