

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Eukaryotic DNA methylation is performed by DNA-methyltransferases that catalyze transfer of a methyl group from S-adenosyl-l-methionine to carbon 5 of cytosine bases in DNA, giving rise to 5-methylcytosine (5-meC). Cytosine methylation is used as an epigenetic mark for maintenance of gene silencing across cellular divisions. However, this chemically stable modification may be removed from DNA through demethylation. DNA demethylation may take place as a passive process due to lack of maintenance methylation during several cycles of DNA replication, or as an active mechanism in the absence of replication. Extensive demethylation of the mammalian genome occurs in preimplantation embryos, first in the male pronucleus through an active mechanism independent of DNA replication and subsequently in both paternal and maternal chromosomes through a passive process. Localized demethylation at specific genes takes place later throughout development and tissue differentiation and rapid cycles of DNA methylation and demethylation of CG dinucleotides at gene promoters have been recently reported. Despite many attempts to identify the mechanism responsible for active DNA demethylation in animal cells, its enzymatic basis remains controversial, although there is evidence for a role of thymine-DNA glycosylase after deamination of 5-meC to thymine. In plants, genetic and biochemical studies have revealed that the Arabidopsis DNA glycosylase domain-containing proteins DME and ROS1 initiate DNA demethylation. Both DME and ROS1 catalyze the release of 5-meC from DNA by a glycosylase/lyase mechanism, cleaving the phosphodiester backbone at the 5-meC removal site by successive β,δ-elimination and leaving a gap that has to be further processed to generate a 3′-OH terminus suitable for polymerization and ligation. This repair-like pathway provides a mechanism to exchange methylated cytosines with cytosines.
Introduction
DNA methylation is found in the genomes of diverse organisms including both prokaryotes and eukaryotes. In prokaryotes, DNA methylation occurs on both cytosine and adenine bases and encompasses part of the host restriction system.1 However, only adenine methylation is used as an epigenetic signal in bacteria, regulating DNA-protein interactions.2 In multicellular eukaryotes methylation seems to be confined to cytosine bases and is associated with an inhibition of gene expression.3,4 Eukaryotic DNA methylation is detected in protists, fungi, plants and animals5 and plays important roles in the establishment of developmental programs6,7 and in genome defense against parasitic mobile elements.8 Hypermethylation of tumour suppressor genes is considered as an important mechanism in the development of many common forms of cancer.9
DNA methylation is performed by DNA-methyltransferases that catalyze transfer of a methyl group from S-adenosyl-l-methionine to cytosine bases in DNA.10 Most of mammalian and plant DNA methylation is restricted to symmetrical CG sequences, but plants also have significant levels of cytosine methylation in the symmetric context CHG (where H is A, C or T) and even in asymmetric sequences.4,11 DNA methylation patterns are established by de novo DNA methyltransferases acting on unmethylated double stranded DNA. Methylation in symmetrical sequences is preserved though cycles of DNA replication by maintenance DNA methyltransferases, which show a preference for hemimethylated substrates and methylate cytosines in the newly synthesized strand.10 Maintenance mechanisms for asymmetric methylation patterns are unknown, but they must include de novo methylation after each cell division.12
There are two general mechanisms by which DNA methylation inhibits gene expression: first, modification of cytosine bases can directly repress transcription by blocking transcriptional activators from binding to cognate DNA sequences;13 and second, proteins that recognize methylated DNA may recruit corepressors to silence gene expression.14-16 There is ample evidence that in vertebrates and plants methyl-CpG-binding proteins (MBPs) use transcriptional corepressor molecules to silence transcription and to modify surrounding chromatin, providing a link between DNA methylation and chromatin remodelling and modification.17-20
DNA methylation is a stable mark for the initiation, establishment and maintenance of gene silencing across cellular divisions, but resembles other macromolecular modifications in being reversible. Demethylation may take place as a passive process due to lack of maintenance methylation during several cycles of DNA replication, or as an active mechanism in the absence of replication.21 In contrast to the well studied genetics, biochemistry and biology of cytosine-DNA-methyltransferases, the enzymatic basis of active demethylation has remained elusive. In this chapter we will review the evidence available on DNA methylation changes in animals and plants and the efforts taken to identify and characterize enzymatic mechanisms of active DNA demethylation. We will see that in animals these mechanisms are still poorly understood, while in plants there is strong genetic and biochemical evidence supporting a base excision process for active DNA demethylation.
Changes in DNA Methylation Patterns in Animals
Although 5-meC is a stable epigenetic mark, DNA methylation patterns are dynamic during animal development, both at the global and local levels. One of the best examples of global changes in DNA methylation takes place in mammalian fertilized oocytes, where methylation is rapidly lost from the paternal genome before the onset of DNA replication.22,23 This active demethylation process is detectable both by immunofluorescence and bisulfite sequencing and does not affect the maternal genome, which is subsequently demethylated through a passive process along cleavage stages. This implies that soon after fertilization paternal sequences are exposed to a putative demethylation machinery while maternal loci are somehow protected from this process.22 Following global demethylation of both parental genomes in preimplantation embryos, mammalian blastocysts undergo a de novo DNA methylation ultimately responsible for the methylation patterns characteristic of the adult animal.24
Genome-wide epigenetic reprogramming through active and/or passive DNA demethylation in zygotes is essential for normal development, as reflected by the difficulty of cloning mammals by somatic cell nuclear transfer (SCNT). In SCNT a differentiated somatic nucleus needs to be reprogrammed in an enucleated oocyte.25 DNA demethylation is absolutely necessary for the reprogramming of somatic cell nuclei,26 but is usually aberrant in mammalian cloned embryos and may contribute to the low efficiency of cloning.27 In fact, it has been shown that the methylation state of the donor nucleus is a major factor governing the efficiency of reprogramming after SCNT.28
Following global demethylation in zygotes, a second genome-wide demethylation wave takes place during germ-cell development. This process has been particularly well studied in mouse embryos, where the primordial germ cells (PGCs) undergo a dramatic and apparently active demethylation process soon after they migrate to the developing gonads.29 This epigenetic reprogramming ensures the erasure of genomic imprints and is critical for returning PGCs to a totipotency state.30
In addition to genome-wide global demethylation, local demethylation occurs in tissue-specific genes throughout development and cellular differentiation.31 In fact, it has been proposed that the methylation pattern generated during development is regulated mainly through demethylation events.24 There is a large body of experimental evidence of local DNA demethylation required for the tissue-specific transcriptional activation of many genes. Some of the best data come from studies with differentiating muscle cells. Demethylation of the α-actin gene is essential to activate its transcription in myoblasts32 and there is a strong correlation between the temporal dynamics of demethylation of a sole CG site of the 5'-flanking region and myogenin expression during muscle differentiation.33 Local demethylation processes are also important for regulation of the immune response: the murine interleukin-2 gene promoter-enhancer region has been shown to be demethylated during T-cell activation34 and demethylation of a unique CG site in its human counterpart is a key memory mark in this regulatory event.35 The IFN-γ promoter is rapidly demethylated upon reestimulation of memory T-cells, but not in naive cells.36 Transcriptional activation by nuclear receptors is also accompanied by methylation changes in the target genes. Thus, glucocorticoid hormones induce stable DNA demethylation within a key enhancer of the rat liver-specific tyrosine aminotransferase (Tat) gene.37 This demethylation is required for the recruitment of additional transcription factors and enhanced transcription activation of the gene. Recent evidence points towards an important role for changes in DNA methylation during memory-associated transcriptional regulation in the nervous system. Thus, demethylation and transcriptional activation of the synaptic plasticity gene reelin occurs in the adult rat hippocampus during fear-conditioning and may be a key process during memory consolidation.38
Changes in DNA Methylation Patterns in Plants
In contrast to mammals, there is no evidence that plants undergo global genome demethylation after fertilization and remethylation during embryo development. On the contrary, there are hints that the DNA methylation status may be stably transmitted across generations. For example, a significantly reduced DNA methylation caused by the ddm1 mutation in Arabidopsis is not restored to normal levels even in a wild-type DDM1 background.39 The absence of a global DNA methylation resetting may have important consequences, given the characteristics of the plant life cycle. Unlike animals, plants do no set aside a dedicated germline early in development and therefore there is a chance that methylation changes in somatic cells are transmitted to the next generation. Thus, hypermethylated epialleles of the Arabidopsis SUPERMAN gene are stable across many generations40 and a naturally occurring hypermethylated version of the LycC gene in Linaria vulgaris is stably inherited.41
Although plants apparently do not experience genome-wide methylation changes, there is some evidence of modifications in DNA methylation during normal plant development. DNA methylation levels in tomato are lower in immature tissues, compared to mature tissues42 and a similar pattern is observed in young seedlings compared to adult rice plants.43 A trend towards increasing DNA methylation during plant development has been also reported in Arabidopsis.44 Surely the local changes in DNA methylation during plant development will receive increased attention with the recent advent of whole genome methylation profiling techniques.45
There is also accumulating evidence of stress-induced changes in DNA methylation patterns. In maize, a significant decrease in DNA methylation is observed in roots upon cold treatment.46 This demethylation was genome-wide but not randomly distributed and since chilled tissues immediately ceased DNA replication it has been suggested to be the result of an active process.46 A specific demethylated region could be identified as a retrotransposon-like sequence designated as ZmMI146 and this is in agreement with the cold-induced DNA demethylation of the Ac/Ds transposon region observed in the same species.47 Activation of transposons in response to stress is well documented in plants and could contribute to genome adaptation to environmental challenges.48 However, demethylation in response to stress is not limited to transposon sequences. In tobacco, a gene encoding a glycerophosphodiesterase-like protein (NtGPDL) and a pathogen-responsive gene (NtAlix1) are demethylated and expressed in response to aluminium stress49 and tobacco mosaic virus infection,50 respectively.
The most compelling evidence of alteration in DNA methylation patterns during plant development comes from imprinted genes. Imprinting is the differential expression of paternal and maternal alleles and has evolved both in placental mammals and flowering plants.51 Imprinting in plants affects the expression of genes in the endosperm.52 The paternal alleles of the Arabidopsis genes MEA, FIS2 and FWA are hypermethylated in the endosperm, whereas the maternal alleles are hypomethylated.53-55 In mammals, maternal-specific expression is achieved by paternal-specific methylation and silencing. In plants, however, methylation is the default state and the maternal expression is attained through the expression in the central cell of DEMETER (DME),55,56 which encodes a DNA glycosylase that excises 5-methylcytosine from DNA56,57 (see below).
The Search for an Enzymatic Mechanism of Active DNA Demethylation in Animal Cells
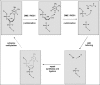
Despite many attempts to identify the mechanism responsible for active DNA demethylation in animal cells, its enzymatic basis remains controversial.21,58 Four major mechanisms have been proposed, according to the initial target of the process and the chemistry involved (Fig. 1).

In the initial search for DNA demethylation mechanisms much attention focused on the identification of a “true” DNA demethylase, i.e., an enzymatic activity that directly removes the methyl group from 5-methylcytosine (Fig. 1A). The first report of an active demethylation mechanism in mammals described an activity that released tritium-labelled methyl-groups from DNA in murine erythroleukemic cells,59 but no further analysis has followed. A DNA demethylase activity that catalyzes the cleavage of a methyl residue from 5-methyl cytosine and releases it as methanol was purified from human cells.60 The same group latter proposed that this activity is identical to MBD2b (methyl CpG-binding domain protein 2b) after testing for demethylase activity following in vitro translation of mRNA derived from the corresponding cDNA.61 However, the demethylating activity of MBD2b could not be independently reproduced by other laboratories19,62 and its relationship to the activity originally described in human cell extracts remains unclear. Furthermore, paternal demethylation in fertilized oocytes lacking MBD2b takes place normally.63 Doubts have also arisen about the viability of the mechanism itself, which involves a thermodynamically unfavourable breakage of the carbon-carbon bond.64 A recent report suggests that the original methanol detection was an artifact and proposes instead that MBD2b catalyzes an oxidative DNA demethylation in which the methyl group is first oxidised to hydroxymethyl and then removed as formaldehyde.65
Other demethylation mechanisms requiring less-challenging chemistry have been described. Studying rat myoblasts, Weiss et al66 proposed that demethylation takes place through the excision of the methylated CG di-nucleotide (Fig. 1B) and suggested a role for RNA since the activity was sensitive to RNAse treatment. The activity was also reported in an independent study67 that reevaluated the role of RNA, suggesting that the inhibitory effects of RNAse were likely due to coating of the DNA substrate. No further characterization of this enzymatic mechanism has been described. Recently, Barreto et al68 reported that the protein Gadd4a (growth arrest and DNA-damage-inducible protein 45 alpha) has a key role in active DNA demethylation in conjunction with XPG, a nuclease that participates in nucleotide excision repair. These authors described that the expression of Gadd45a activates methylation-silenced reported plasmids, promotes gene-specific and global demethylation and is required to avoid hypermethylation in cultured human cells.68 However, these results have been challenged by Jin et al,69 who argue that Gadd45a is not expressed in oocytes and zygotes, as would be expected from a demethylation factor. Jin et al were unable to substantiate a role of Gadd45a in DNA demethylation69 and casted doubts about the quantification of 5-meC in cells after knockdown of Gadd45a reported by Barreto et al.68 Therefore, the possible role of Gad45a in DNA demethylation through nucleotide excision is uncertain.
A third proposed mechanism for active demethylation is removal of 5-meC as a free base followed by replacement with an unmodified cytosine70 (Fig. 1C). The initial step of this mechanism would be catalyzed by a DNA glycosylase that cleaves the labile N-glycosidic bond between the 5-meC base and the deoxyribose moiety in DNA, leaving an abasic site that must be further processed. DNA glycosylases initiate the base excision repair (BER) pathway, which in most organisms removes common base modifications (oxidation, deamination, alkylation) caused by endogenous genotoxic agents.71 A 5-methylcytosine-DNA glycosylase activity that releases 5-meC from DNA was identified and partially purified from human cells.72,73 A similar activity identified in chicken embryos74 was found to copurify with a protein homologous to human thymine DNA glycosylase (TDG).75,76 It was latter reported that methyl CpG binding protein 4 (MBD4), another human DNA glycosylase with no sequence similarity to TDG, also has 5-methylcytosine-DNA glycosylase activity.77 Both TDG and MBD4 are DNA glycosylases that show a preference for U and T in U·G and T·G mismatches located within a CG context.78,79 However, they have been shown to have a very weak activity on 5-meC in 5-meC·G pairs compared to their activities towards U·G and T·G mismatches77,79,80 and hence their precise roles in DNA demethylation remain unclear.
Although the exact mechanism for DNA demethylation in mammals is still unknown, the evidence in favour of a DNA repair-based process seems to accumulate. It has been reported that the demethylation upstream the tyrosine aminotransferase (Tat) gene upon activation by the glucocorticoid receptor occurs independently of DNA replication and involves the generation of nicks in the DNA backbone 3′ to the 5-meC.81 In addition, DNA demethylation in mouse PGCs occurs before histone replacement, which supports a DNA-repair based mechanism.82
The notion that DNA demethylation may involve a DNA-repair process has fuelled the search for plausible mechanisms performed by known enzymes. One of the leading hypothesis is that demethylation might be achieved indirectly though deamination of 5-meC and repair of the ensuing T·G mismatch by a DNA glycosylase83 (Fig. 1D). The enzymes Aid and Apobec1 have been put forward as candidate deaminases in this process, since they both have 5-meC deaminase activity.84 Furthermore, Aid and Apobec1 genes are colocalized within a cluster of pluripotency genes and are expressed in oocytes and primordial germ cells, which undergo epigenetic reprogramming.84 Since deaminases require single-stranded DNA, the initiation of DNA demethylation would probably need accessory proteins, such as chromatin remodeling and/or transcription factors, to expose 5-meC residues to deaminase action.84
The idea that demethylation may be initiated by a deamination process has received unexpectedly strong support from recent work with the ligand-dependent transcription factor estrogen receptor α (ERα).85,86 ERα induces a cyclical activation of its targets promoters through ordered and periodic recruitment of a series of coactivator complexes, defining a “transcriptional clock” that limits the transcriptional response.87 Two recent studies report that DNA methylation shows a similar cyclical pattern of demethylation and remethylation at the promoter of the ERα-responsive gene pS285 and four other target genes.86 Remarkably, DNA methyltransferases Dnmt3a and b are cyclically recruited to the pS2 promoter at the beginning of each transcriptionally productive cycle, when demethylation occurs, together TDG and other proteins that participate in BER.85 TDG is required for DNA demethylation and transcriptional activation of the pS2 promoter, as demonstrated though reduction of its expression by short interfering RNA (siRNA).85 Dnmt3a and b can deaminate 5-meC to thymine in vitro in the absence of the methyl donor S-adenosyl methionine (SAM), generating T·G mismatches that are substrates for TDG.85 The authors propose that the rapid DNA demethylation observed during the transcriptional cycles ERα-responsive genes is achieved though the repair of T·G mismatches arising from 5-meC deamination catalyzed by Dnmt3a and b.85 This model implies that Dnmts are involved both in DNA methylation and demethylation. Although the model is attractive, additional data about the efficiency of deamination activity of Dnmts in vivo are needed.
Active DNA Demethylation in Plants
In plants, there is convincing genetic and biochemical evidence that a family of DNA glycosylase domain-containing proteins typified by Arabidopsis DME (DEMETER) and ROS1 (REPRESSOR OF SILENCING 1) initiate erasure of 5-meC through a base excision repair process. DME was identified in a search for mutations causing parent-of-origin effects on seed viability88 and is expressed primarily in the central cell of the female gametophyte, where it is required for the expression of the maternal alleles of the imprinted genes MEA, FWA and FIS2.54,55,88 In the case of MEA imprinting, mutations in the methyltransferase gene MET1 suppress the requirement for DME53 and the maternal MEA allele is not hypomethylated in dme endosperm.56 ROS1 was identified in a screen for mutants with deregulated expression of the repetitive RD29A-LUC transgene.89 Whereas in wild plants the transgene and the homologous endogenous gene are expressed, ros1 mutants display transcriptional silencing and hypermethylation of both loci.89
In addition to DME and ROS1, the genome of Arabidopsis encodes two additional paralogs, referred to as DEMETER-LIKE proteins DML2 and DML3.88 All four proteins are large polypeptides containing a DNA glycosylase domain with significant sequence similarity to base excision DNA repair proteins in the HhH-GPD superfamily, named after its hallmark helix-hairpin-helix and Gly/Pro rich loop followed by a conserved aspartate.90 The HhH motif present in DME, ROS1, DML2 and DML3 includes an invariant lysine conserved in the subset of DNA glycosylases/lyases, able both to hydrolyze the N-glycosidic bond linking bases to DNA and to cleave the phosphodiester backbone at the site where a base has been removed.91 In addition to the DNA glycosylase domain, the proteins of the DME/ROS1 family share two other conserved domains of unknown function.57 The HhH-GPD superfamily of DNA glycosylases is widespread in all three domains of life (bacteria, archea and eukaryotes) and its members are typically 200-400 amino acids long.92 However, proteins of the DME/ROS1 family are unusually large (1100-2000 amino acids) compared to typical DNA glycosylases. Furthermore, they appear to be unique to plants, with putative orthologs present in mosses and unicellular green algae. This suggests that active demethylation though excision of 5-meC may have appeared early during plant evolution.
DME and ROS1 are the best characterized in vitro among the members of this family of atypical DNA glycosylases (Fig. 2).56,57,93 Both DME and ROS1 remove 5-meC as a free base from DNA through a glycosylase/lyase mechanism57 and cleave the phosphodiester backbone at the 5-meC removal site by successive β,δ-elimination, leaving a gap that has to be further processed to generate a 3'-OH terminus suitable for polymerization and ligation.56,57,93 Excision of 5-meC in vitro is more efficient on those sequences more likely to be methylated in vivo. Thus, DME and ROS1 erase 5-meC at CG, CHG and CHH sequences, with a preference for CG sites56,57 which matches the pattern of DNA methylation in plants. Furthermore, both proteins remove 5-meC more efficiently from a CAG context than when located in the outer position of a CCG context,57 in agreement with the fact that CCG is the sequence showing the lowest methylation level among CHG sites.94 DML2 and DML3 are also 5-meC DNA glycosylases/lyases.95,96 While DML2 activity is very weak, at least in vitro, DML3 shows an enzymatic activity comparable to those of DME and ROS1, with a similar substrate specificity.95,96 In addition to 5-meC paired to guanine, DME, ROS1 and DML3 also remove thymine from a T·G mismatch located at CG, CHG and CHH sequences.56,57,96 Therefore, it cannot be ruled out the possibility that DME, ROS1 and/or DML3 also play a role in neutralizing the mutagenic consequences of the spontaneous deamination of 5-meC to thymine through their activity on T·G mismatches. The effect of dme and ros1 mutations on mutagenesis in vivo has not been assessed, but could be compounded by their epigenetic effects on plant development.
As noted above, an active demethylation pathway initiated by TDG and/or MBD4 DNA glycosylases acting on 5-meC has been proposed in animal cells.75,77,97,98 However, it has been argued that the main in vivo role for both proteins is to counteract the mutagenic potential of 5-meC and C deamination in CG sequences,79,99 given their high efficiency on U·G and T·G mismatches,76,78,79 compared to their weak activity on 5-meC·G base pairs.77,80 The proteins of the DME/ROS1 family are structurally unrelated to TDG, which belongs to a large group of uracil-DNA glycosylases different from the HhH-GPD family,100 but share with MBD4 a HhH-GPD DNA glycosylase domain located at the C-terminal half of the protein. However, unlike MBD4, they do not have a methyl-CpG binding domain.79 In contrast to the strong substrate specificity of TDG and MBD4 for T·G and U·G mismatches, DME and ROS1 show a preference for 5-meC over a T·G mismatch in a CG sequence context, the most frequent DNA methylation target in plant and animal genomes and they do not display detectable activity on U·G mispairs.57 Thus, the biochemical properties of DME and ROS1, together with the available genetic evidence, suggest that an important role for both enzymes in vivo is excision of 5-meC.
It remains to be explained how these enzymes locate and recognize 5-meC in DNA. The methylated cytosine is not a “lesion”, such as other base modifications from endogenous or exogenous origin that are substrates of repair DNA glycosylases. However, the extent of the 5-meC influence on DNA structure is largely unknown although may alter its hydration pattern.101 An understanding of how plant 5-meC DNA glycosylases specifically recognize their target base will require solving their crystal structure in complex with DNA.
The precise in vivo roles of plant 5-meC DNA glycosylases are not fully understood. DME is probably required to specifically initiate erasure of 5-meC at MEA, FWA, FIS2 and perhaps other unidentified loci, in female gametes before fertilization.54-56 ROS1 is needed to prevent transcriptional gene silencing and hypermethylation of a repetitive transgene, but the observation of developmental abnormalities in ros1 mutants after inbreeding89 suggests that it also regulates expression of endogenous loci. In fact, CHG and CHH sites become hypermethylated at FWA and several transposons in ros1 mutants, with an additional slight increase in CG methylation.102 Furthermore, microarray analysis allowed the identification of several genes with reduced expression in ros1 plants and some of these showed hypermethylation at their promoter regions.102 A recent report based on 5-meC immunocapturing followed by genome-tiling microarrays analysis described the identification of about 200 regions that become hypermethylated in a ros1 dml2 dml3 triple mutant.95 Most of the hypermethylation was located at genic regions, did not affect any particular gene class and accumulated predominantly at the 5′and 3′ends of genes, which is opposite to the methylation pattern found in wild-type plants.95 Much of the DNA methylation in Arabidopsis is directed by RNA interference (RNAi) pathways and the hypermethylated regions observed in ros1 dml2 dml3 mutants are enriched for small interfering RNAs.103 Furthermore, there is genetic evidence that ROS1 demethylation antagonizes de novo methylation directed by different RNAi pathways.103 Altogether, these results suggest that an important in vivo function for ROS1, DML2 and DML3 is to protect the genome from excess methylation.
By other hand, a detailed analysis of the methylation distribution at the FWA gene and AtGP1 transposon in wild-type and dml mutant plants suggests that DML2 and DML3 may play additional roles in methylation dynamics.96 Mutations in DML2 and/or DML3 lead to hypermethylation of cytosine residues that are unmethylated or weakly methylated in wild-type plants, in agreement with a role in protecting the genome from excess methylation. But, intriguingly, sites that are heavily methylated in wild-type plants are hypomethylated in mutants.96 Furthermore, a recent report describing the analysis of the Arabidopsis methylome at single-base resolution found, as expected, hundreds of discrete hypermethylated regions in a ros1 dml2 dml3 triple mutant, but also sites where the methylation levels were lower than in the wild-type.104 Altogether, these results suggest that ROS1, DML2 and DML3 are required not only for removing DNA methylation marks from improperly-methylated cytosines, but also for maintenance of high methylation levels in properly targeted sites.
Conclusions and Future Prospects
It is somewhat paradoxical that the mechanism of active DNA demethylation in animals, where evidence of global and local demethylation is abundant, is less well understood than in plants (see also in the chapter by Faines et al and by Parisien and Bhagwat in this volume). There is ample evidence supporting a DNA repair-based process during active DNA demethylation in mammals, but the responsible mechanism(s) is (are) still unknown. Recent data support a model in which demethylation is initiated by 5-meC deamination, followed by thymine excision from the ensuing T·G mismatch and replacement with an unmodified cytosine. When considering a repair-mediated DNA mechanism, it should be contemplated the possibility that the enzymes that carry out the deamination step in local and global demethylation processes may be different. While Dnmt3a and b may initiate the rapid demethylation observed during cyclical transcriptional activation, cytidine deaminases such as Aid and Apobec1 have the catalytic activity and expression pattern adequate to initiate the paternal global demethylation observed in zygotes. There is some evidence that TDG may be responsible for the T·G repair step during local demethylation at transcriptionally active promoters, but its role, if any, during global demethylation remains unknown. In this regard, it is important to remark that tdg-null mouse embryos die during mid-gestation, while other DNA glycosylases, including another thymine-DNA glycosylase such as MBD4, are dispensable for embryonic development.105 However, no study on the capacity of tdg-null zygotes to perform DNA demethylation has been reported so far.
While the evidence available in animal cells remains fragmentary, our current understanding of DNA demethylation in plants is more solid but still far from complete. The data available about 5-meC DNA glycosylases of the DME/ROS1 family indicates that plant cells are able of use base excision not only to remove lesions from DNA but also to erasure naturally occurring modified residues. The emerging notion is that an important role of 5-meC glycosylases is to protect the genome from excess methylation and this is in agreement with their likely evolution from ancient enzymes dedicated to genome maintenance. It remains to be determined how this protective role fits with the specific function of DME in activating the maternal alleles of imprinted genes. It is possible that plants have availed themselves of the differential expression of a 5-meC DNA glycosylase such as DME in male and female gametophytes for control of imprinting. In addition, recent data suggest that ROS1, DML2 and DML3 may be required not only to protect from deleterious methylation but also to maintain high methylation levels at appropriately targeted sites. Therefore, the final scenario for DNA demethylation in plants may be more complex than previously suspected, with dynamic DNA methylation/demethylation processes contributing both to the stability and flexibility of the epigenome.
Acknowledgements
Work in our laboratory is supported by grants from the Ministerio de Educación y Ciencia, Spain and the Junta de Andalucía, Spain.
References
- 1.
- Wilson GG, Murray NE. Restriction and modification systems. Annu Rev Genet. 1991;25:585–627. [PubMed: 1812816]
- 2.
- Casadesus J, Low D. Epigenetic gene regulation in the bacterial world. Microbiol Mol Biol Rev. 2006;70(3):830–856. [PMC free article: PMC1594586] [PubMed: 16959970]
- 3.
- Bender J. DNA methylation and epigenetics. Annu Rev Plant Physiol Plant Mol Biol. 2004;55:41–68. [PubMed: 15725056]
- 4.
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002;16(1):6–21. [PubMed: 11782440]
- 5.
- Colot V, Rossignol JL. Eukaryotic DNA methylation as an evolutionary device. Bioessays. 1999;21(5):402–411. [PubMed: 10376011]
- 6.
- Holliday R, Pugh JE. DNA modification mechanisms and gene activity during development. Science. 1975;187(4173):226–232. [PubMed: 1111098]
- 7.
- Riggs AD. X inactivation, differentiation and DNA methylation. Cytogenet Cell Genet. 1975;14(1):9–25. [PubMed: 1093816]
- 8.
- Yoder JA, Walsh CP, Bestor TH. Cytosine methylation and the ecology of intragenomic parasites. Trends Genet. 1997;13(8):335–340. [PubMed: 9260521]
- 9.
- Esteller M. Aberrant DNA methylation as a cancer-inducing mechanism. Annu Rev Pharmacol Toxicol. 2005;45:629–656. [PubMed: 15822191]
- 10.
- Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases. Annu Rev Biochem. 2005;74:481–514. [PubMed: 15952895]
- 11.
- Finnegan EJ, Genger RK, Peacock WJ. et al. DNA methylation in plants. Annu Rev Plant Physiol Plant Mol Biol. 1998;49:223–247. [PubMed: 15012234]
- 12.
- Finnegan EJ, Kovac KA. Plant DNA methyltransferases. Plant Mol Biol. 2000;43(2-3):189–201. [PubMed: 10999404]
- 13.
- Watt F, Molloy PL. Cytosine methylation prevents binding to DNA of a HeLa cell transcription factor required for optimal expression of the adenovirus major late promoter. Genes Dev. 1988;2(9):1136–1143. [PubMed: 3192075]
- 14.
- Boyes J, Bird A. DNA methylation inhibits transcription indirectly via a methyl-CpG binding protein. Cell. 1991;64(6):1123–1134. [PubMed: 2004419]
- 15.
- Hendrich B, Bird A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol. 1998;18(11):6538–6547. [PMC free article: PMC109239] [PubMed: 9774669]
- 16.
- Hark AT, Schoenherr CJ, Katz DJ. et al. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature. 2000;405(6785):486–489. [PubMed: 10839547]
- 17.
- Jones PL, Veenstra GJ, Wade PA. et al. Methylated DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet. 1998;19(2):187–191. [PubMed: 9620779]
- 18.
- Nan X, Ng HH, Johnson CA. et al. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature. 1998;393(6683):386–389. [PubMed: 9620804]
- 19.
- Ng HH, Zhang Y, Hendrich B. et al. MBD2 is a transcriptional repressor belonging to the MeCP1 histone deacetylase complex. Nat Genet. 1999;23(1):58–61. [PubMed: 10471499]
- 20.
- Zemach A, Grafi G. Characterization of Arabidopsis thaliana methyl-CpG-binding domain (MBD) proteins. Plant J. 2003;34(5):565–572. [PubMed: 12787239]
- 21.
- Kress C, Thomassin H, Grange T. Local DNA demethylation in vertebrates: how could it be performed and targeted? FEBS Lett. 2001;494(3):135–140. [PubMed: 11311228]
- 22.
- Oswald J, Engemann S, Lane N. et al. Active demethylation of the paternal genome in the mouse zygote. Curr Biol. 2000;10(8):475–478. [PubMed: 10801417]
- 23.
- Mayer W, Niveleau A, Walter J. et al. Demethylation of the zygotic paternal genome. Nature. 2000;403(6769):501–502. [PubMed: 10676950]
- 24.
- Weiss A, Cedar H. The role of DNA demethylation during development. Genes Cells. 1997;2(8):481–486. [PubMed: 9348038]
- 25.
- Cibelli JB. et al. Principles of cloning. Amsterdam: Academic Press. 2002
- 26.
- Simonsson S, Gurdon J. DNA demethylation is necessary for the epigenetic reprogramming of somatic cell nuclei. Nat Cell Biol. 2004;6(10):984–990. [PubMed: 15448701]
- 27.
- Dean W, Santos F, Stojkovic M. et al. Conservation of methylation reprogramming in mammalian development: aberrant reprogramming in cloned embryos. Proc Natl Acad Sci USA. 2001;98(24):13734–13738. [PMC free article: PMC61110] [PubMed: 11717434]
- 28.
- Blelloch R, Wang Z, Meissner A. et al. Reprogramming efficiency following somatic cell nuclear transfer is influenced by the differentiation and methylation state of the donor nucleus. Stem Cells. 2006;24(9):2007–2013. [PMC free article: PMC3000431] [PubMed: 16709876]
- 29.
- Hajkova P, Erhardt S, Lane N. et al. Epigenetic reprogramming in mouse primordial germ cells. Mech Dev. 2002;117(1-2):15–23. [PubMed: 12204247]
- 30.
- Surani MA, Hayashi K, Hajkova P. Genetic and epigenetic regulators of pluripotency. Cell. 2007;128(4):747–762. [PubMed: 17320511]
- 31.
- Frank D, Keshet I, Shani M. et al. Demethylation of CpG islands in embryonic cells. Nature. 1991;351(6323):239–241. [PubMed: 2041571]
- 32.
- Paroush Z, Keshet I, Yisraeli J. et al. Dynamics of demethylation and activation of the alpha-actin gene in myoblasts. Cell. 1990;63(6):1229–1237. [PubMed: 2261641]
- 33.
- Lucarelli M, Fuso A, Strom R. et al. The dynamics of myogenin site-specific demethylation is strongly correlated with its expression and with muscle differentiation. J Biol Chem. 2001;276(10):7500–7506. [PubMed: 11096088]
- 34.
- Bruniquel D, Schwartz RH. Selective, stable demethylation of the interleukin-2 gene enhances transcription by an active process. Nat Immunol. 2003;4(3):235–240. [PubMed: 12548284]
- 35.
- Murayama A, Sakura K, Nakama M. et al. A specific CpG site demethylation in the human interleukin 2 gene promoter is an epigenetic memory. EMBO J. 2006;25(5):1081–1092. [PMC free article: PMC1409718] [PubMed: 16498406]
- 36.
- Kersh EN, Fitzpatrick DR, Murali-Krishna K. et al. Rapid demethylation of the IFN-gamma gene occurs in memory but not naive CD8 T-cells. J Immunol. 2006;176(7):4083–4093. [PubMed: 16547244]
- 37.
- Thomassin H, Flavin M, Espinas ML. et al. Glucocorticoid-induced DNA demethylation and gene memory during development. EMBO J. 2001;20(8):1974–1983. [PMC free article: PMC125428] [PubMed: 11296230]
- 38.
- Miller CA, Sweatt JD. Covalent modification of DNA regulates memory formation. Neuron. 2007;53(6):857–869. [PubMed: 17359920]
- 39.
- Kakutani T, Munakata K, Richards EJ. et al. Meiotically and mitotically stable inheritance of DNA hypomethylation induced by ddm1 mutation of Arabidopsis thaliana. Genetics. 1999;151(2):831–838. [PMC free article: PMC1460490] [PubMed: 9927473]
- 40.
- Jacobsen SE, Meyerowitz EM. Hypermethylated SUPERMAN epigenetic alleles in arabidopsis. Science. 1997;277(5329):1100–1103. [PubMed: 9262479]
- 41.
- Cubas P, Vincent C, Coen E. An epigenetic mutation responsible for natural variation in floral symmetry. Nature. 1999;401(6749):157–161. [PubMed: 10490023]
- 42.
- Messeguer R, Ganal MW, Steffens JC. et al. Characterization of the level, target sites and inheritance of cytosine methylation in tomato nuclear DNA. Plant Mol Biol. 1991;16(5):753–770. [PubMed: 1859863]
- 43.
- Sha AH, Lin XH, Huang JB. et al. Analysis of DNA methylation related to rice adult plant resistance to bacterial blight based on methylation-sensitive AFLP (MSAP) analysis. Mol Genet Genomics. 2005;273(6):484–490. [PubMed: 15968537]
- 44.
- Ruiz-Garcia L, Cervera MT, Martinez-Zapater JM. et al. DNA methylation increases throughout Arabidopsis development. Planta. 2005;222(2):301–306. [PubMed: 15968513]
- 45.
- Zhu JK. Epigenome sequencing comes of age. Cell. 2008;133(3):395–397. [PMC free article: PMC3137521] [PubMed: 18455978]
- 46.
- Steward N, Ito M, Yamaguchi Y. et al. Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress. J Biol Chem. 2002;277(40):37741–37746. [PubMed: 12124387]
- 47.
- Steward N, Kusano T, Sano H. Expression of ZmMET1, a gene encoding a DNA methyltransferase from maize, is associated not only with DNA replication in actively proliferating cells, but also with altered DNA methylation status in cold-stressed quiescent cells. Nucleic Acids Res. 2000;28(17):3250–3259. [PMC free article: PMC110715] [PubMed: 10954592]
- 48.
- Wessler SR. Turned on by stress. Plant retrotransposons. Curr Biol. 1996;6(8):959–961. [PubMed: 8805314]
- 49.
- Choi CS, Sano H. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol Genet Genomics. 2007;277(5):589–600. [PubMed: 17273870]
- 50.
- Wada Y, Miyamoto K, Kusano T. et al. Association between up-regulation of stress-responsive genes and hypomethylation of genomic DNA in tobacco plants. Mol Genet Genomics. 2004;271(6):658–666. [PubMed: 15148604]
- 51.
- Feil R, Berger F. Convergent evolution of genomic imprinting in plants and mammals. Trends Genet. 2007;23(4):192–199. [PubMed: 17316885]
- 52.
- Huh JH, Bauer MJ, Hsieh TF. et al. Endosperm gene imprinting and seed development. Curr Opin Genet Dev. 2007;17(6):480–485. [PMC free article: PMC2180190] [PubMed: 17962010]
- 53.
- Xiao W, Gehring M, Choi Y. et al. Imprinting of the MEA Polycomb gene is controlled by antagonism between MET1 methyltransferase and DME glycosylase. Dev Cell. 2003;5(6):891–901. [PubMed: 14667411]
- 54.
- Jullien PE, Kinoshita T, Ohad N. et al. Maintenance of DNA methylation during the Arabidopsis life cycle is essential for parental imprinting. Plant Cell. 2006;18(6):1360–1372. [PMC free article: PMC1475502] [PubMed: 16648367]
- 55.
- Kinoshita T, Miura A, Choi Y. et al. One-way control of FWA imprinting in Arabidopsis endosperm by DNA methylation. Science. 2004;303(5657):521–523. [PubMed: 14631047]
- 56.
- Gehring M, Huh JH, Hsieh TF. et al. DEMETER DNA glycosylase establishes MEDEA polycomb gene self-imprinting by allele-specific demethylation. Cell. 2006;124(3):495–506. [PMC free article: PMC4106368] [PubMed: 16469697]
- 57.
- Morales-Ruiz T, Ortega-Galisteo AP, Ponferrada-Marin MI. et al. DEMETER and REPRESSOR OF SILENCING 1 encode 5-methylcytosine DNA glycosylases. Proc Natl Acad Sci USA. 2006;103(18):6853–6858. [PMC free article: PMC1458983] [PubMed: 16624880]
- 58.
- Wolffe AP, Jones PL, Wade PA. DNA demethylation. Proc Natl Acad Sci USA. 1999;96(11):5894–5896. [PMC free article: PMC34201] [PubMed: 10339513]
- 59.
- Gjerset RA, Martin DW Jr. Presence of a DNA demethylating activity in the nucleus of murine erythroleukemic cells. J Biol Chem. 1982;257(15):8581–8583. [PubMed: 7047522]
- 60.
- Ramchandani S, Bhattacharya SK, Cervoni N. et al. DNA methylation is a reversible biological signal. Proc Natl Acad Sci USA. 1999;96(11):6107–6112. [PMC free article: PMC26843] [PubMed: 10339549]
- 61.
- Bhattacharya SK, Ramchandani S, Cervoni N. et al. A mammalian protein with specific demethylase activity for mCpG DNA. Nature. 1999;397(6720):579–583. [PubMed: 10050851]
- 62.
- Wade PA, Gegonne A, Jones PL. et al. Mi-2 complex couples DNA methylation to chromatin remodelling and histone deacetylation. Nat Genet. 1999;23(1):62–66. [PubMed: 10471500]
- 63.
- Santos F, Hendrich B, Reik W. et al. Dynamic reprogramming of DNA methylation in the early mouse embryo. Dev Biol. 2002;241(1):172–182. [PubMed: 11784103]
- 64.
- Smith SS. Gilbert's conjecture: the search for DNA (cytosine-5) demethylases and the emergence of new functions for eukaryotic DNA (cytosine-5) methyltransferases. J Mol Biol. 2000;302(1):1–7. [PubMed: 10964556]
- 65.
- Hamm S, Just G, Lacoste N. et al. On the mechanism of demethylation of 5-methylcytosine in DNA. Bioorg Med Chem Lett. 2008;18(3):1046–1049. [PubMed: 18162397]
- 66.
- Weiss A, Keshet I, Razin A. et al. DNA demethylation in vitro: involvement of RNA. Cell. 1996;86(5):709–718. [PubMed: 8797818]
- 67.
- Swisher JF, Rand E, Cedar H. et al. Analysis of putative RNase sensitivity and protease insensitivity of demethylation activity in extracts from rat myoblasts. Nucleic Acids Res. 1998;26(24):5573–5580. [PMC free article: PMC148033] [PubMed: 9837985]
- 68.
- Barreto G, Schafer A, Marhold J. et al. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature. 2007;445(7128):671–675. [PubMed: 17268471]
- 69.
- Jin SG, Guo C, Pfeifer GP. GADD45A does not promote DNA demethylation. PLoS Genet. 2008;4(3):e1000013. [PMC free article: PMC2265528] [PubMed: 18369439]
- 70.
- Razin A, Szyf M, Kafri T. et al. Replacement of 5-methylcytosine by cytosine: a possible mechanism for transient DNA demethylation during differentiation. Proc Natl Acad Sci USA. 1986;83(9):2827–2831. [PMC free article: PMC323399] [PubMed: 3458243]
- 71.
- Lindahl T, Wood RD. Quality control by DNA repair. Science. 1999;286(5446):1897–1905. [PubMed: 10583946]
- 72.
- Vairapandi M, Duker NJ. Enzymic removal of 5-methylcytosine from DNA by a human DNA-glycosylase. Nucleic Acids Res. 1993;21(23):5323–5327. [PMC free article: PMC310565] [PubMed: 8265344]
- 73.
- Vairapandi M, Duker NJ. Partial purification and characterization of human 5-methylcytosine-DNA glycosylase. Oncogene. 1996;13(5):933–938. [PubMed: 8806682]
- 74.
- Jost JP, Siegmann M, Sun L. et al. Mechanisms of DNA demethylation in chicken embryos. Purification and properties of a 5-methylcytosine-DNA glycosylase. J Biol Chem. 1995;270(17):9734–9739. [PubMed: 7730351]
- 75.
- Zhu B, Zheng Y, Hess D. et al. 5-methylcytosine-DNA glycosylase activity is present in a cloned G/T mismatch DNA glycosylase associated with the chicken embryo DNA demethylation complex. Proc Natl Acad Sci USA. 2000;97(10):5135–5139. [PMC free article: PMC25794] [PubMed: 10779566]
- 76.
- Neddermann P, Gallinari P, Lettieri T. et al. Cloning and expression of human G/T mismatch-specific thymine-DNA glycosylase. J Biol Chem. 1996;271(22):12767–12774. [PubMed: 8662714]
- 77.
- Zhu B, Zheng Y, Angliker H. et al. 5-Methylcytosine DNA glycosylase activity is also present in the human MBD4 (G/T mismatch glycosylase) and in a related avian sequence. Nucleic Acids Res. 2000;28(21):4157–4165. [PMC free article: PMC113156] [PubMed: 11058112]
- 78.
- Sibghat U, Gallinari P, Xu YZ. et al. Base analog and neighboring base effects on substrate specificity of recombinant human G:T mismatch-specific thymine DNA-glycosylase. Biochemistry. 1996;35(39):12926–12932. [PubMed: 8841138]
- 79.
- Hendrich B, Hardeland U, Ng HH. et al. The thymine glycosylase MBD4 can bind to the product of deamination at methylated CpG sites. Nature. 1999;401(6750):301–304. [PubMed: 10499592]
- 80.
- Hardeland U, Bentele M, Jiricny J. et al. The versatile thymine DNA-glycosylase: a comparative characterization of the human, Drosophila and fission yeast orthologs. Nucleic Acids Res. 2003;31(9):2261–2271. [PMC free article: PMC154230] [PubMed: 12711670]
- 81.
- Kress C, Thomassin H, Grange T. Active cytosine demethylation triggered by a nuclear receptor involves DNA strand breaks. Proc Natl Acad Sci USA. 2006;103(30):11112–11117. [PMC free article: PMC1544051] [PubMed: 16840560]
- 82.
- Hajkova P, Ancelin K, Waldmann T. et al. Chromatin dynamics during epigenetic reprogramming in the mouse germ line. Nature. 2008;452(7189):877–881. [PMC free article: PMC3847605] [PubMed: 18354397]
- 83.
- Reik W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature. 2007;447(7143):425–432. [PubMed: 17522676]
- 84.
- Morgan HD, Dean W, Coker HA. et al. Activation-induced cytidine deaminase deaminates 5-methylcytosine in DNA and is expressed in pluripotent tissues: implications for epigenetic reprogramming. J Biol Chem. 2004;279(50):52353–52360. [PubMed: 15448152]
- 85.
- Metivier R, Gallais R, Tiffoche C. et al. Cyclical DNA methylation of a transcriptionally active promoter. Nature. 2008;452(7183):45–50. [PubMed: 18322525]
- 86.
- Kangaspeska S, Stride B, Metivier R. et al. Transient cyclical methylation of promoter DNA. Nature. 2008;452(7183):112–115. [PubMed: 18322535]
- 87.
- Metivier R, Penot G, Hubner MR. et al. Estrogen receptor-alpha directs ordered, cyclical and combinatorial recruitment of cofactors on a natural target promoter. Cell. 2003;115(6):751–763. [PubMed: 14675539]
- 88.
- Choi Y, Gehring M, Johnson L. et al. DEMETER, a DNA glycosylase domain protein, Is required for endosperm gene imprinting and seed viability in Arabidopsis. Cell. 2002;110(1):33–42. [PubMed: 12150995]
- 89.
- Gong Z, Morales-Ruiz T, Ariza RR. et al. ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell. 2002;111(6):803–814. [PubMed: 12526807]
- 90.
- Nash HM, Bruner SD, Scharer OD. et al. Cloning of a yeast 8-oxoguanine DNA glycosylase reveals the existence of a base-excision DNA-repair protein superfamily. Curr Biol. 1996;6(8):968–980. [PubMed: 8805338]
- 91.
- Krokan HE, Standal R, Slupphaug G. DNA glycosylases in the base excision repair of DNA. Biochem J. 1997;325:1–16. [PMC free article: PMC1218522] [PubMed: 9224623]
- 92.
- Denver DR, Swenson SL, Lynch M. An evolutionary analysis of the helix-hairpin-helix superfamily of DNA repair glycosylases. Mol Biol Evol. 2003;20(10):1603–1611. [PubMed: 12832627]
- 93.
- Agius F, Kapoor A, Zhu JK. Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation. Proc Natl Acad Sci USA. 2006;103(31):11796–11801. [PMC free article: PMC1544249] [PubMed: 16864782]
- 94.
- Cokus SJ, Feng S, Zhang X. et al. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature. 2008;452(7184):215–219. [PMC free article: PMC2377394] [PubMed: 18278030]
- 95.
- Penterman J, Zilberman D, Huh JH. et al. DNA demethylation in the Arabidopsis genome. Proc Natl Acad Sci USA. 2007;104:6752–6757. [PMC free article: PMC1847597] [PubMed: 17409185]
- 96.
- Ortega-Galisteo AP, Morales-Ruiz T, Ariza RR. et al. Arabidopsis DEMETER-LIKE proteins DML2 and DML3 are required for appropriate distribution of DNA methylation marks. Plant Mol Biol. 2008;67(6):671–681. [PubMed: 18493721]
- 97.
- Jost JP, Oakeley EJ, Zhu B. et al. 5-Methylcytosine DNA glycosylase participates in the genome-wide loss of DNA methylation occurring during mouse myoblast differentiation. Nucleic Acids Res. 2001;29(21):4452–4461. [PMC free article: PMC60186] [PubMed: 11691933]
- 98.
- Zhu B, Benjamin D, Zheng Y. et al. Overexpression of 5-methylcytosine DNA glycosylase in human embryonic kidney cells EcR293 demethylates the promoter of a hormone-regulated reporter gene. Proc Natl Acad Sci USA. 2001;98(9):5031–5036. [PMC free article: PMC33158] [PubMed: 11296268]
- 99.
- Barnes DE, Lindahl T. Repair and genetic consequences of endogenous DNA base damage in mammalian cells. Annu Rev Genet. 2004;38:445–476. [PubMed: 15568983]
- 100.
- Aravind L, Koonin EV. The alpha/beta fold uracil DNA glycosylases: a common origin with diverse fates Genome Biol 20001(1) research0007.0001-0007.0008 . [PMC free article: PMC15025] [PubMed: 11178247]
- 101.
- Marcourt L, Cordier C, Couesnon T. et al. Impact of C5-cytosine methylation on the solution structure of d(GAAAACGTTTTC)2. An NMR and molecular modelling investigation. Eur J Biochem. 1999;265(3):1032–1042. [PubMed: 10518799]
- 102.
- Zhu J, Kapoor A, Sridhar VV. et al. The DNA glycosylase/lyase ROS1 functions in pruning DNA methylation patterns in Arabidopsis. Curr Biol. 2007;17(1):54–59. [PubMed: 17208187]
- 103.
- Penterman J, Uzawa R, Fischer RL. Genetic interactions between DNA demethylation and methylation in Arabidopsis. Plant Physiol. 2007;145(4):1549–1557. [PMC free article: PMC2151691] [PubMed: 17951456]
- 104.
- Lister R, O'Malley RC, Tonti-Filippini J. et al. Highly integrated single-base resolution maps of the epigenome in Arabidopsis. Cell. 2008;133(3):523–536. [PMC free article: PMC2723732] [PubMed: 18423832]
- 105.
- Cortazar D, Kunz C, Saito Y. et al. The enigmatic thymine DNA glycosylase. DNA Repair (Amst). 2006;6(4):489–504. [PubMed: 17116428]
- 106.
- Morgan HD, Santos F, Green K. et al. Epigenetic reprogramming in mammals. Hum Mol Genet. 2005;(14 Spec No 1):R47–58. [PubMed: 15809273]
- DNA Demethylation - Madame Curie Bioscience DatabaseDNA Demethylation - Madame Curie Bioscience Database
- Role of Mammalian Coronin 7 in the Biosynthetic Pathway - Madame Curie Bioscienc...Role of Mammalian Coronin 7 in the Biosynthetic Pathway - Madame Curie Bioscience Database
- Lysosomal Storage Disorders - Madame Curie Bioscience DatabaseLysosomal Storage Disorders - Madame Curie Bioscience Database
- Neural Crest Delamination and Migration: Integrating Regulations of Cell Interac...Neural Crest Delamination and Migration: Integrating Regulations of Cell Interactions, Locomotion, Survival and Fate - Madame Curie Bioscience Database
- Dynamic Connections of Nuclear Envelope Proteins to Chromatin and the Nuclear Ma...Dynamic Connections of Nuclear Envelope Proteins to Chromatin and the Nuclear Matrix - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...