

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

Composition
As in other animals, the nuclear envelope in Drosophila has three basic subparts: the nuclear membranes, a lamina just beneath the membrane layer, and nuclear pores that rivet the nuclear membranes and permit nucleo-cytoplasmic transport.
There are two nuclear membranes, each a lipid bilayer containing characteristic proteins. The outer nuclear membrane is continuous with the endoplasmic reticulum. The inner nuclear membrane contains proteins that can associate with lamin and/or chromatin. The Drosophila genome predicts genes for relatives of several proteins known to be in the inner membranes of vertebrates or nematodes. These include otefin, a LEM protein,5-7 which has been shown to be a constituent of the Drosophila inner nuclear membrane, and predicted counterparts of two other LEM proteins (emerin and MAN-1) as well as nurim, the lamin B receptor (LBR), and unc-84 (see ref. 7); we still await reports of the subcellular localization of these predicted proteins.
The Drosophila nuclear lamina contains proteins related to those in other animal nuclear laminas. In particular, its major constituent is intermediate-filament proteins of the lamin family. As in most higher animals, Drosophila has lamins of two subtypes: A-type and B-type. However, its suite of lamins is simpler than that in vertebrates. Drosophila has a single gene for a B-type lamin called lamin Dm0.8 Lamin Dm0 derivatives (called, collectively, lamin Dm, below) are present in the nuclear envelopes of nearly all Drosophila cells9–14 and, this protein is essential for full viability.15 Fertility of Drosophila females also demands sufficient levels of lamin Dm0 derivatives in the female germ line.15 The B-type lamin undergoes posttranslational modifications, including phosphorylations that convert it from a soluble form (Dmmit) to nuclear forms that can make polymeric fibers in the lamina (Dm1, Dm2).11,16Drosophila's single A-type lamin (“lamin C”,17–19) analogous to the vertebrate situation, is found only in certain cell types; in Drosophila these are a subset of differentiated cells. Drosophila nuclear laminas in early embryos include at least one protein without a vertebrate counterpart—the Young Arrest (“YA”) protein,1,20 a novel hydrophilic nuclear lamina component that is essential for embryonic development to initiate (see below). Made only during oogenesis (though excluded from nuclei at this time21,22), YA is found only in the nuclei of young (cleavage stage) embryos;1,20 after this time it is not detected in any Drosophila cells, except for developing oocytes.
Drosophila nuclear envelopes also contain pore complexes whose components are relatives of those of vertebrate nuclear pores:23 including nup154,24 gp210,25 a Tpr homologue,26 myosin-like proteins27,28 and a predicted nup153 homologue with RNA binding activity.29 Components of the nuclear import/export system, including importin family members and an exportin homologue, have also been found in Drosophila (e.g., see refs. 30-39).
Cell cycle dynamics: In at least some, and possibly all,Drosophila cells, nuclear envelopes do not completely break down during mitosis. This was first shown for cleavage stage Drosophila embryos, in an elegant electron microscopy study;40 subsequent investigators extended this finding by documenting the behaviors of specific nuclear envelope, or envelope-associated, proteins during the cell cycles in embryos.1,5,26 The envelopes of interphase nuclei in embryos are continuous around the nuclei. By prometaphase, the envelopes are seen to be open at the spindle poles. A second membrane layer, paralleling the nuclear membranes and, like them, open at the spindle poles, begins to form around the original nuclear envelope. Nuclear pores dissociate from the nuclear membranes beginning in prophase.26,40 In metaphase, the nuclear lamina largely dissociates: most of its lamin Dm disappears from the nuclear periphery5 and YA also becomes undetectable at the nuclear periphery.1 However, the fenestrated, double “spindle envelope” remains, still open at the poles. Chromosomes segregate on the spindle within this envelope. At telophase, lamin Dm, otefin and nuclear pore proteins begin to be detected in the nuclear envelopes, and the second membrane layer disappears.1,5,26,40 At the end of telophase in early embryos, the YA protein becomes detectable at the nuclear periphery, suggesting that it assembles into the nuclear envelope after lamin Dm.1 These nuclear envelope dynamics may be characteristic of all Drosophila nuclei: spindle envelopes of similar appearance to those reported by Stafstrom and Staehelin40 have also been seen in spermatogenic cells41and in cultured Drosophila cells.42
The essential role of the nuclear envelope in basic cellular events such as DNA replication, nuclear organization, gene expression and the cell cycle is well known in many organisms. Recently it has become clear that the nuclear envelope plays an important role in development and in particular cell types (e.g., 1-4). The stages surrounding the formation of pronuclei in fertilized eggs involve dramatic changes in nuclear envelope composition, organization and function, making them of particular interest for investigating nuclear envelope dynamics. Knowing the basis of the changes in nuclear envelopes as one goes from gamete to differentiating embryo is instructive both to our understanding of nuclear assembly/disassembly and function, and to our understanding of this critical developmental stage. This article focuses on pronuclear and embryo nuclear envelope formation and function in Drosophila melanogaster. I will first describe the overall dynamics and composition of Drosophila nuclear envelopes, and then their developmental dynamics in the stages surrounding pronuclear formation and early embryonic development.
Developmental Changes in Nuclear Envelopes Around the Time of Fertilization
Dramatic changes in nuclear envelopes occur around the time of fertilization in animals. At these times, highly differentiated gametes, with unique nuclear structures, combine to form a zygote. The gamete nuclei are converted to pronuclei that then participate in a special series of events that generates the first zygotic nuclei of the embryo. In many animals, including Drosophila, the zygotic nuclei then go through a very rapid mitotic stage (cleavage), using maternal proteins stored in the egg. When the maternal “dowry” has been depleted, the zygotic genome takes control of cell division dynamics, and usually at about the same time the zygotic cells begin to undertake differentiative pathways.
The next sections of this article follow these events in Drosophila. I briefly describe the formation of gametes, pronuclei, zygotic nuclei, cleavage nuclei and nuclei committed to differentiative pathways. I then focus on the composition and changes in nuclear envelopes that accompany these events. If the roles of nuclear envelopes or their constituents are known, these are included. As illustrated in Fig. 1, some nuclear envelope components are nearly ubiquitous throughout these stages, others are present in, and essential for, the functions of particular nuclei, and still others appear or disappear when nuclei have certain characteristics or fates.
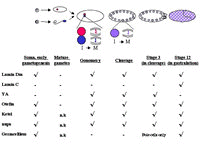
Gamete Nuclei
Oocytes
The development of an oocyte begins with 4 mitotic divisions by the daughter of a germ line stem cell (see ref. 43 for review); these divisions have incomplete cytokinesis. Fifteen of the cells produced in these divisions (the “nurse cells”) then cease dividing. They endoreplicate their genomes and begin to synthesize RNAs and proteins that will be transferred to the oocyte to provision it for subsequent embryo development. The 16th cell, the oocyte, initiates meiosis, arresting in metaphase of meiosis I at the end of oogenesis. This arrest is maintained until the oocyte is ovulated.44 Ovulation activates the oocyte to complete meiosis; though meiotic progression occurs at the time of fertilization, it is independent of sperm penetration.44–47 Meiosis I and, immediately thereafter, meiosis II occur (for reviews see refs. 47-50). The process is extremely rapid, taking as little as 17 min to complete. It occurs without cytokinesis, and results in the presence of 4 haploid meiotic products in the activated egg.
Several nuclear envelope proteins have unique behaviors, or biochemical properties, in developing or activating oocytes. Oocyte nuclei (and nurse cell nuclei) initially have standard nuclear envelope compositions, including lamin Dm,11 otefin51 and nup154.24 By late oogenesis, the distribution of these proteins in the oocyte nucleus appears to be different from that in typical nuclei (including the nurse cell nuclei): higher levels of lamin Dm and otefin are detected within the oocyte nucleus (relative to levels at its periphery) than in other cell types.11,51 Whether this reflects a difference in nuclear structure, leading to a greater accessibility of internal lamin Dm to staining or is due to a redistribution of lamin Dm at this stage is not known. In late-stage oocytes, there is also a soluble form of lamin Dm, that remains detectable until the end of the maternally-driven cleavage-cycle phase of embryonic development.11 This lamin isoform, which is presumably the lamin Dm donated to the oocyte by the nurse cells and stored for use in embryo mitosis, is thought to be the same as the mitotic isoform of lamin, Dmmit, that is observed in tissue culture cells.11 Finally, the YA protein is detected in oocytes; this is the first time it is found postembryonically. However, this YA is in a highly phosphorylated form that is excluded from the nuclei, apparently by associating with a cytoplasmic retention complex.21,22
At the very end of oogenesis, the metaphase-arrested nucleus of the oocyte appears to disassemble: lamin Dm and otefin become undetectable at the oocyte's nuclear periphery, though the precise structure of any remaining nuclear envelope is not known.11,51 The dynamics of the nuclear envelope during the rapid meiotic divisions that follow are also unknown. The interphase between meiosis I and meiosis II is extremely short and hard to “catch”, and there may be no typical interphase at all. In preliminary experiments we have been unable to detect assembly of a nuclear lamina around the meiotic nuclei during the time between telophase of meiosis I and metaphase of meiosis II.52
The nuclear envelope plays important roles in oogenesis. Females homozygous for a partial loss-of-function allele of the lamin Dm0 gene are sterile, indicating the essential function of this lamin in oogenesis.15At least part of the function of lamin Dm may relate to its role in polarity generation in the oocyte. In normal oocytes, the nucleus is located at the anterior end of the cell, and is closer to one side of the oocyte (see ref. 53 for review). That side is fated to become the dorsal side of the egg and embryo. mRNA from the gurken (grk) gene (a TGF-alpha family member) accumulates near the oocyte nucleus (see ref. 53 for review). This results in higher levels of GRK protein in the vicinity of the oocyte nucleus, which in turn activates a signaling system that confers a dorsal identity on the side of the oocyte nearest the nucleus. In oocytes carrying misguided mutations of the lamin Dm0 gene, grk RNA is mis-localized, and the oocyte's dorsoventral polarity is consequently disrupted.54 Thus, the nuclear envelope contributes to proper positioning of dorsoventral determinants in the developing oocyte.
Sperm
The development of sperm begins with mitotic divisions by the daughter of a stem cell in the male germline (see ref. 55 for review). After 4 mitotic divisions with incomplete cytokinesis, the nuclei undertake meiosis. After meiosis has completed, spermiogenesis occurs. During this process, the sperm nucleus becomes elongated and bounded by microtubules and the sperm itself elongates and loses most of its cytoplasm. There are dramatic changes in the nuclear envelope of spermatogenic cells during this process. The nuclei in the cells undergoing mitosis have nuclear envelopes that contain lamin Dm,13,56 otefin51 and nup154,24and can accommodate YA if it is ectopically expressed in these cells.13 The distribution of other nuclear envelope components in spermatogenic cells has not been reported. The nuclear envelopes of spermatogenic cells have spindle envelopes, and cell cycle dynamics of the type described above.41 During spermiogenesis, when the sperm nucleus' shape changes, the composition of its nuclear envelope also changes. All detectable nuclear envelope antigens tested (lamin Dm, otefin, and, upon ectopic expression, YA13,51,56) become undetectable around the sperm nuclei. Membranes are reported to remain around the sperm nucleus at least until late in this stage. The nucleus of a mature Drosophila sperm either does not have a nuclear envelope or is bounded by an atypical envelope whose protein components are not cross-reactive with any somatic nuclear envelope protein tested thus far.
Pronuclei
Female
After the oocyte nucleus has completed meiosis, nuclear envelopes form around the four female meiotic products. All four meiotic products are within the egg cytoplasm and are indistinguishable in terms of nuclear envelope antigens at this time, even though their fates will be different. The haploid nucleus closest to the center of the egg will usually become the female pronucleus and contribute to the embryo's genome. The other three meiotic products will migrate to the egg periphery, where these “polar bodies” will enter a metaphase-like state but then degenerate (see ref. 48 for review).
The envelopes of the four products of oocyte meiosis contain lamin Dm and the YA protein. The presence of lamin Dm in the envelopes of these nuclei likely is simply a reflection of the return to standard nuclear envelope structure. YA's presence in the envelopes of the postmeiotic nuclei marks, however, the initial appearance of this protein at the nuclear periphery YA was produced, but excluded from nuclei, during oogenesis.21 YA's phosphorylation level drops during egg activation.21 We hypothesize that this releases YA from a cytoplasmic retention complex, and allows it to enter nuclei for the first time when nuclei form after meiosis.22
Male
If the egg is penetrated by a sperm, the sperm's nucleus will have to be converted to a male pronucleus in order to participate in development. As in systems that are presently better characterized (e.g., see refs. 57,58 for review), the sperm nucleus' chromatin must decondense (presumably involving replacement of some of its chromatin-packaging proteins with proteins available in the egg), change shape,13,14 and acquire a new nuclear envelope made from maternally provided constituents including lamin Dm and YA.13 Since the entire Drosophila sperm enters the egg, still bounded by its plasma membrane,59 the first step in converting the sperm nucleus to a male pronucleus is removal of the sperm's plasma membrane. The product of the sneaky (snky) gene appears to be necessary for this membrane removal; evidence suggests that sperm from males mutant in snky fail to demembranate when they enter eggs.60 In eggs fertilized by snky sperm, the sperm nucleus fails to decondense, change shape or migrate, and it does not become surrounded by lamin Dm or YA.13,60
Once the sperm nucleus has been exposed to the cytoplasm of the fertilized egg, it begins to swell and decondense its DNA. Partway through the decondensation process, the sperm nucleus acquires an envelope made from components present in the egg.13,14,61 Analysis of subsequent steps in Drosophila male pronuclear formation have been most informed by genetic analysis, since existing in vitro nuclear assembly systems62–64 have not been ideal for detailed examination of nuclear assembly: these systems are too inefficient, perhaps because they are made from embryos at mixed stages of the cell cycle.63 The in vitro assembly systems have, however, been useful for identifying chromatin decondensation factors that may act in pronuclear formation.65,66
Phenotypic analysis of mutants in the maternal-effect gene sésame (ssm)14 suggest that Drosophila male pronuclear formation follows the paradigm elegantly established in sea urchins, in which decondensation of the sperm nucleus occurs in two phases: an initial decondensation independent of acquisition of a new nuclear envelope, and a later decondensation dependent on a nuclear envelope derived from maternal components (see refs. 57,58,67,68 for review). In eggs lacking functional ssm gene product, sperm nuclei decondense only partially. The partially decondensed sperm nuclei have acquired nuclear envelopes that include lamin Dm and YA (provided by the egg).14 The phenotype of the sperm nuclei in ssm embryos suggests that ssm may encode a maternal factor needed for the second, nuclear envelope-dependent, phase of male pronuclear formation. The molecular nature of the ssm gene product has not yet been reported.
Combining the Pronuclear Genomes
Once the sperm nucleus has acquired a maternally provided nuclear envelope, microtubules from its associated centriole capture one of the female meiotic products (the female pronucleus). The female and male pronuclei, each bounded by a lamin Dm- and YA-containing nuclear envelope, migrate into the anterior/center of the embryo and become closely apposed, but do not fuse1, 61, 69 (see ref. 48 for review). The apposed pronuclei then initiate the first cell cycle of the early embryo. Pronuclear DNA replicates. A single spindle forms, using centrosomes that derived from a centriole donated by the sperm. A metaphase occurs, with the maternal chromosomes remain on one side of the spindle and the paternal ones on the other. The two chromosome complements then undergo anaphase, in coordination but again each on their own side of the spindle.1,61,69 These characteristics lead to the designation of this division as “gonomeric”. In late telophase of the gonomeric division, the parental chromosome complements finally mix; thus the first zygotic nuclei form at the end of the first cell cycle. Two independent lines of evidence confirm the light-microscopy determination of gonomery. In eggs of normal females fertilized by sperm from Wolbachia-infected male flies, development arrests soon after fertilization.70,71 Many such zygotes show abnormalities during the gonomeric division. Their male-derived chromosomes are abnormally condensed and confined to one side of the gonomeric spindle; the maternal genome segregates, as normal, on the other side of the spindle.72The phenotype of the ssm mutant also shows that maternal and paternal chromosomes segregate separately during the gonomeric division. In fertilized ssm eggs, one side of the spindle is occupied by condensed male chromatin that cannot participate in the division, but maternally derived chromatids still segregate as normal on the other side of the spindle.14
In these developmental events as well, the nuclear envelope and its components play important roles. Function of the YA protein is essential for the nuclei to enter the gonomeric cell cycle. In the absence of YA function, the apposed pronuclei appear to arrest prior to S phase of this cell cycle1,73,74 Our data lead us to hypothesize that YA function is necessary for nuclei to pass a checkpoint that monitors completion of meiosis and creation of mitotically competent nuclei. YA is capable of binding to chromatin, via interaction with DNA and histone H2B,63,75 and also interacts with lamin Dm0.76We hypothesize that its function is to confer a mitotically competent state on nuclei.74 Another nuclear envelope-related protein whose function is apparent at this early time in development is Ketel, a Drosophila homologue of importin-β.34 Ketel protein is made during oogenesis and placed into eggs; it continues to be made from the zygotic genome in embryos.34 Embryos lacking maternal Ketel product, or embryos produced by females carrying apparent dominant-negative Ketel mutations arrest development during the gonomeric cell cycle, with nuclei that appear abnormal. The arrest phenotype of these embryos indicates that Ketel is required for proper nuclear assembly following mitosis.34,35 The mechanism by which Ketel exerts this function is not known. The simplest possible model—that Ketel mutations prevent all nuclear import and hence assembly of nuclei with proper nuclear envelopes—is ruled out since at least one substrate (cNLS-phycoerythrin) can enter nuclei in the mutant embryos. This suggests that Ketel has either a unique function specific to nuclear assembly at the end of mitosis77,78 or that the import of at least one molecule essential for nuclear assembly is completely dependent on Ketel, with no alternative pathway available for its import.34,35
The nuclear envelope also confers the “gonomeric” character on this first cell cycle. Immunofluorescence studies showed that the separation of the parental genomes during the gonomeric division is due to the presence of a spindle envelope around each of the pronuclei.61 The spindle envelopes, open at the poles, allow microtubules of the single spindle to penetrate, but keep the parental chromosome separate until they reach the spindle poles late in telophase.61
Cleavage Nuclei
The gonomeric division is followed by 12 very rapid mitotic divisions (reviewed in ref. 48), leading to several thousand nuclei. These divisions occur within a syncytium, are roughly synchronous, and are driven by maternal products that were stored in the egg during oogenesis. After about 7 nuclear cycles (including the gonomeric division) the nuclei begin to migrate to the periphery of the egg, forming a “syncytial blastoderm” (see ref. 48 for review). Towards the end of the syncytial phase of development, the division cycles slow from ∼9 min to ∼21 min, as maternal products are depleted. In the final cleavage cell cycle, the maternally provided mRNA for string (stg), which encodes a cdc25-family phosphatase, is degraded. Further progress through mitotic cycles requires transcription of stg from the zygotic genome, and accumulation of STG protein to sufficient levels.79–81 At this time also, membranes grow down into the egg, separating each nucleus into its own cell (initiating the cellular blastoderm stage). In such cellularized embryos, mitotic cycles become more independent, losing the synchrony seen in syncytial blastoderm embryos; however nuclei of cells with similar fate often share similar cell division behaviors (mitotic domains; see ref. 82).
During the precellularization nuclear division cycles, nuclear envelopes show typical Drosophila nuclear envelope dynamics,40 involving cyclic loss, and then restoration of nuclear pores and their associated proteins, otefin and lamin Dm (in telophase),1,5,26,40 YA (later in telophase),1 and presence of fenestrated membranes and a spindle envelope during mitosis. Lamin Dm0 derivatives show changes in phosphorylation status that are thought to correspond to the appearance of nuclear, polymerized forms (Dm1, Dm2) in addition to the soluble form (thought to be Dmmit) that was already present in early embryos.11
The nuclear envelope has several essential developmental functions during the cleavage stages. First, the timing of nuclear envelope reassembly plays an important role in cell cycle dynamics at this stage. A checkpoint, identified through the phenotype of the grapes (grp) mutation, involves the role of the nuclear envelope.83,84 grp mutants arrest because their nuclei undertake cell cycles with improperly condensed chromatin. Careful examination of the dynamics of a variety of nuclear structures in grp vs. normal embryos in the presence and absence of aphidicolin, indicated that the defect in grp embryos results in premature breakdown of nuclear envelopes in those embryos. This premature breakdown occurs before the chromosomes are fully condensed or fully replicated, resulting in abnormal chromosome behaviors during mitosis.84 The GRP protein is normally located in nuclei, but is released when nuclear envelopes break down; it is suggested to be part of a checkpoint that prevents nuclear envelope breakdown until S-phase is completed,84 also allowing for attainment of the proper condensation state by chromosomes. A second role for the nuclear envelope is suggested by the tight parallel of lamin's, and its position and dynamics to those of mitotic spindles.85 It was proposed that the nuclear lamina might play a role in facilitating spindle formation or dynamics during mitosis.85 A third role for the nuclear envelope is more indirect, in the generation of some embryo polarities during this developmental stage. This role results from the nuclear envelope acting as a barrier that separates nuclear contents from the cytoplasm: dorsoventral polarity, for example, is controlled by the regulated access of the DL transcription factor to the nuclear interior. On the dorsal side of the embryo, a cytoplasmic complex keeps DL out of nuclei, causing them to take on a dorsal fate (e.g., see ref. 86 for review). Disruption of this complex on the ventral side of the embryo due to a signaling pathway (e.g., see ref. 87 for review) allows DL to enter nuclei on this side, conferring on them a ventral fate. Though roles, if any, for specific nuclear envelope proteins in the regulation of dorsoventral polarity are not known, this example serves to illustrate the critical role of a functional nuclear envelope in developmental choices.
Nuclei of Differentiating Cells in Embryos
The first overt differentiative change in the embryo is detected in the behavior of ∼8 nuclei in an otherwise cleavage-stage embryo. During cycle 9 (counting the gonomeric division) these most posterior nuclei of the embryo bud out into individual cells. These “pole cells” are fated to form the germ line. Their divisional timing differs from the synchronous syncytial divisions of the remaining somatic nuclei in the embryo. The nuclear pores of pole cell nuclear envelopes acquire an associated protein that is the product of the germcellless (gcl) gene.88,89Function of gcl is essential for pole cell nuclei to take on a germ cell fate. gcl is also expressed at other times in development, but its function at those times is not known. A vertebrate counterpart of gcl has recently been reported to be involved in regulating transcription,90 suggesting that modulation of transcription by GCL protein in pole cells may be important in specifying their fate.
Other nuclei and cells in Drosophila embryos only show signs of commitment to a differentiated fate after the cellular blastoderm stage. As nuclei take on differentiated fates, the composition of their envelopes change from that of cleavage nuclei: YA protein disappears1,20 by the end of the blastoderm stage and later, during gastrulation, lamin C becomes detectable in nuclear envelopes of differentiated cells (initially in the nuclear envelopes of cells that will become oenocytes, hindgut and posterior spiracle, and later in additional cell types19).
The nuclear envelope is again seen to play important roles in the differentiative stages. For example, misguided mutant alleles of the lamin Dm0 gene have a phenotype in differentiating cells: these mutations specifically disrupt the branching growth of trachae that occurs late in embryogenesis.54 The aberrant growth and targeting by the tracheal branches suggests that lamin Dm plays a role in cell polarity.
Conclusion
Nuclear envelopes undergo dramatic changes in composition and dynamics during the developmental transition from gametes to differentiating embryos. These changes include modification and partial dissolution in gametogenesis, complete reformation from maternal components in early embryos, and modulation of composition during embryogenesis as nuclei take on differentiation competence. In Drosophila, we now know much about the dynamics and functions of nuclear envelopes at these critical times, and several of the likely important molecular players. Continued exploitation of the genetics, genomics and molecular developmental biology in this model system will provide a detailed picture of the mechanism by which pronuclear envelopes form, break down, and regulate the start of development.
Acknowledgments
I thank my YA-colleagues past and present for many stimulating discussions, Drs. M. Goldberg, J. Liu and J. Yu for many helpful comments on this manuscript, and the NIH (R01-GM44659) for supporting our work in this area.
References
- 1.
- Lin H, Wolfner MF. The Drosophila maternal-effect gene fs(1)Ya encodes a cell cycle-dependent nuclear envelope component required for embryonic mitosis. Cell. 1991;64:49–62. [PubMed: 1986869]
- 2.
- Furukawa K, Hotta Y. cDNA cloning of a germ cell specific lamin B3 from mouse spermatocytes and analysis of its function by ectopic expression in somatic cells. EMBO J. 1993;12:97–106. [PMC free article: PMC413179] [PubMed: 8094052]
- 3.
- Manilal S, Nguyen TM, Sewry CA. et al. The Emery-Dreifuss muscular dystrophy protein, emerin, is a nuclear membrane protein. Hum Mol Genet. 1996;5:801–808. [PubMed: 8776595]
- 4.
- Bonne G, Di Barletta MR, Varnous S. et al. Mutations in the gene encoding lamin A/C cause autosomal dominant EmeryDreifuss muscular dystrophy. Nat Genet. 1999;21:285–288. [PubMed: 10080180]
- 5.
- Harel A, Zlotkin E, Nainudel-Epszteyn S. et al. Persistence of major nuclear envelope antigens in an envelopelike structure during mitosis in Drosophila melanogaster embryos. J Cell Sci. 1989;94:463–470. [PubMed: 2517292]
- 6.
- Padan R, NainudelEpszteyn S, Goitein R. et al. Isolation and characterization of the Drosophila nuclear envelope otefin cDNA. J Biol Chem. 1990;265:7808–7813. [PubMed: 2186029]
- 7.
- Cohen M, Lee KK, Wilson KL. et al. Transcriptional repression, apoptosis, human disease and the functional evolution of the nuclear lamina. Trends Biochem Sci. 2001;26:41–47. [PubMed: 11165516]
- 8.
- Gruenbaum Y, Landesman Y, Drees B. et al. Drosophila nuclear lamin precursor Dm0 is translated from either of two developmentally regulated mRNA species apparently encoded by a single gene. J Cell Biol. 1988;106:585–596. [PMC free article: PMC2115108] [PubMed: 3126192]
- 9.
- Smith DE, Fisher PA. Identification, developmental regulation, and response to heat shock of two antigenically related forms of a major nuclear envelope protein in Drosophila embryos: application of an improved method for affinity purification of antibodies using polypeptides immobilized on nitrocellulose blots. J Cell Biol. 1984;99:20–28. [PMC free article: PMC2275611] [PubMed: 6203917]
- 10.
- Smith DE, Gruenbaum Y, Berrios M. et al. Biosynthesis and interconversion of Drosophila nuclear lamin isoforms during normal growth and in response to heat shock. J Cell Biol. 1987;105:771–790. [PMC free article: PMC2114747] [PubMed: 3624309]
- 11.
- Smith DE, Fisher PA. Interconversion of Drosophilanuclear lamin isoforms during oogenesis, early embryogenesis, and upon entry of cultured cells into mitosis. J Cell Biol. 1989;108:255–265. [PMC free article: PMC2115415] [PubMed: 2492999]
- 12.
- Stuurman N, Maus N, Fisher PA. Interphase phosphorylation of the Drosophila nuclear lamin: sitemapping using a monoclonal antibody. J Cell Sci. 1995;108:3137–3144. [PubMed: 8537453]
- 13.
- Liu J, Lin H, Lopez JM. et al. Formation of the male pronuclear lamina in Drosophila. Dev Biol. 1997;184:187–196. [PubMed: 9133429]
- 14.
- Loppin B, Docquier M, Bonneton F. et al. The maternal-effect mutation sésame affects the formation of the male pronucleus in Drosophila melanogaster. Dev Biol. 2000;222:392–404. [PubMed: 10837127]
- 15.
- LenzBöhme B, Wismar J, Fuchs S. et al. Insertional mutagenesis of the Drosophila nuclear lamin Dm0 gene results in defective nuclear envelopes, clustering of nuclear pore complexes, and accumulation of annulate lamellae. J Cell Biol. 1997;137:1001–1016. [PMC free article: PMC2136230] [PubMed: 9166402]
- 16.
- Schneider U, Mini T, Jeno P. et al. Phosphorylation of the major Drosophila lamin in vivo: site identification during both Mphase (meiosis) and interphase by electrospray ionization tandem mass spectrometry. Biochemistry. 1999;38:4620–4632. [PubMed: 10194384]
- 17.
- Bossie CA, Sanders MM. A cDNA from Drosophila melanogaster encodes a lamin Clike intermediate filament protein. J Cell Sci. 1993;104:1263–1272. [PubMed: 8314904]
- 18.
- Riemer D, Weber K. The organization of the gene for Drosophila lamin C: limited homology with vertebrate lamin genes and lack of homology versus the Drosophila lamin Dmo gene. Eur J Cell Biol. 1994;63:299–306. [PubMed: 8082654]
- 19.
- Riemer D, Stuurman N, Berrios M. et al. Expression of Drosophila lamin C is developmentally regulated: Analogies with vertebrate A-type lamins. J Cell Sci. 1995;108:3189–3198. [PubMed: 7593280]
- 20.
- Lopez J, Song K, Hirshfeld A. et al. The Drosophila fs(1)Ya protein, which is needed for the first mitotic division, is in the nuclear lamina and in the envelopes of cleavage nuclei, pronuclei and nonmitotic nuclei. Dev Biol. 1994;163:202–211. [PubMed: 8174776]
- 21.
- Yu J, Liu J, Song K. et al. Nuclear entry of the Drosophila melanogaster nuclear lamina protein YA correlates with developmentally regulated changes in its phosphorylation state. Dev Biol. 1999;210:124–134. [PubMed: 10364432]
- 22.
- Yu J, Garfinkel A, Wolfner MF. Interaction of the essential Drosophila nuclear protein YA with P0/AP3 in the cytoplasm: implications for developmental regulation of YA's subcellular location. Dev Biol. 2002;244:429–441. [PubMed: 11944949]
- 23.
- Filson AJ, Lewis A, Blobel G. et al. Monoclonal antibodies prepared against the major Drosophila nuclear matrixpore complexlamina glycoprotein bind specifically to the nuclear envelope in situ. J Biol Chem. 1985;260:3164–3172. [PubMed: 3919018]
- 24.
- Gigliotti S, Callaini G, Andone S. et al. Nup154, a new Drosophila gene essential for male and female gametogenesis is related to the nup155 vertebrate nucleoporin gene. J Cell Biol. 1998;142:1195–1207. [PMC free article: PMC2149350] [PubMed: 9732281]
- 25.
- Berrios M, Meller VH, McConnell M. et al. Drosophila gp210, an invertebrate nuclear pore complex glycoprotein. Eur J Cell Biol. 1995;67:1–7. [PubMed: 7641726]
- 26.
- Zimowska G, Aris JP, Paddy MR. A Drosophila Tpr protein homolog is localized both in the extrachromosomal channel network and to nuclear pore complexes. J Cell Sci. 1997;110:927–944. [PubMed: 9152019]
- 27.
- Berrios M, Fisher PA, Matz EC. Localization of a myosin heavy chainlike polypeptide to Drosophila nuclear pore complexes. Proc Natl Acad Sci USA. 1991;88:219–223. [PMC free article: PMC50781] [PubMed: 1986370]
- 28.
- Strambio-DeCastillia C, Blobel G, Rout MP. Proteins connecting the nuclear pore complex with the nuclear interior. J Cell Biol. 1999;144:839–855. [PMC free article: PMC2148185] [PubMed: 10085285]
- 29.
- Dimaano C, Ball JR, Prunuske AJ. et al. RNA association defines a functionally conserved domain in the nuclear pore protein Nup153. J Biol Chem. 2001;276:45349–45357. [PubMed: 11567018]
- 30.
- Torok I, Strand D, Schmitt R. et al. The overgrown hematopoietic organs 31 tumor suppressor gene of Drosophila encodes an importin-like protein accumulating in the nucleus at the onset of mitosis. J Cell Biol. 1995;129:1473–1489. [PMC free article: PMC2291178] [PubMed: 7790349]
- 31.
- Siomi MC, Fromont M, Rain JC. et al. Functional conservation of the transportin nuclear import pathway in divergent organisms. Mol Cell Biol. 1998;18:4141–4148. [PMC free article: PMC108998] [PubMed: 9632798]
- 32.
- Dockendorff TC, Tang Z, Jongens TA. Cloning of karyopherin-alpha3 from Drosophila through its interaction with the nuclear localization sequence of germcellless protein. Biol Chem. 1999;380:1263–1272. [PubMed: 10614818]
- 33.
- Herold A, Suyama M, Rodrigues JP. et al. TAP (NXF1) belongs to a multigene family of putative RNA export factors with a conserved modular architecture. Mol Cell Biol. 2000;20:8996–9008. [PMC free article: PMC86553] [PubMed: 11073998]
- 34.
- Lippai M, Tirian L, Boros I. et al. The Ketel gene encodes a Drosophila homologue of importinbeta. Genetics. 2000;156:1889–1900. [PMC free article: PMC1461400] [PubMed: 11102382]
- 35.
- Tirian L, Puro J, Erdelyi M. et al. The Ketel(D) dominant-negative mutations identify maternal function of the Drosophila importin-beta gene required for cleavage nuclei formation. Genetics. 2000;156:1901–1912. [PMC free article: PMC1461349] [PubMed: 11102383]
- 36.
- Fasken MB, Saunders R, Rosenberg M. et al. A leptomycin Bsensitive homologue of human CRM1 promotes nuclear export of nuclear export sequencecontaining proteins in Drosophila cells. J Biol Chem. 2000;275:1878–1886. [PubMed: 10636888]
- 37.
- Mathe E, Bates H, Huikeshoven H. et al. Importin-alpha3 is required at multiple stages of Drosophila development and has a role in the completion of oogenesis. Dev Biol. 2000;223:307–322. [PubMed: 10882518]
- 38.
- Collier S, Chan HY, Toda T. et al. The Drosophila embargoed gene is required for larval progression and encodes the functional homolog of Schizosaccharomyces Crm1. Genetics. 2000;155:1799–1807. [PMC free article: PMC1461193] [PubMed: 10924475]
- 39.
- Lorenzen JA, Baker SE, Denhez F. et al. Nuclear import of activated DERK by DIM7, an importin family member encoded by the gene moleskin. Development. 2001;128:1403–1414. [PubMed: 11262240]
- 40.
- Stafstrom JP, Staehelin LA. Dynamics of the nuclear envelope and of nuclear pore complexes during mitosis in the Drosophila embryo. Eur J Cell Biol. 1984;34:179–189. [PubMed: 6428889]
- 41.
- Church K, Lin H P P. Meiosis in Drosophila melanogaster: II. The prometaphase1 kinetochore microtubule bundle and kinetochore orientation in males. J Cell Biol. 1982;93:365–373. [PMC free article: PMC2112869] [PubMed: 6807996]
- 42.
- Debec A, Marcaillou C. Structural alterations of the mitotic apparatus induced by the heat shock response in Drosophila cells. Biol Cell. 1997;89:67–78. [PubMed: 9297784]
- 43.
- Spradling AC. Developmental genetics of oogenesis, in The development of Drosophila melanogaster, Bate, M. and Martinez Arias, A., Editors. Cold Spring Harbor Laboratory Press: Plainview, New York: 19931–70.
- 44.
- Heifetz Y, Yu J, Wolfner MF. Ovulation triggers activation of Drosophila oocytes. Dev Biol. 2001;234:416–424. [PubMed: 11397010]
- 45.
- Doane WW. Completion of meiosis in uninseminated eggs of Drosophila melanogaster. Science. 1960;132:677–678. [PubMed: 13817039]
- 46.
- Mahowald AP, Goralski TJ, Caulton JH. In vitro activation of Drosophila eggs. Dev Biol. 1983;98:437–445. [PubMed: 6409691]
- 47.
- Page AW, OrrWeaver TL. Activation of the meiotic divisions in Drosophila oocytes. Dev Biol. 1997;183:195–207. [PubMed: 9126294]
- 48.
- Foe VE, Odell GM, Edgar BA. Mitosis and morphogenesis in the Drosophila embryo: point and counterpoint, in The development of Drosophila melanogaster, Bate, M. and Martinez Arias, A., Editors. Plainview: Cold Spring Harbor Laboratory Press, 1993149–300.
- 49.
- OrrWeaver TL. Meiosis in Drosophila: Seeing is believing. Proc Natl Acad Sci, USA. 1995;92:10443–10449. [PMC free article: PMC40628] [PubMed: 7479817]
- 50.
- Endow SA, Komma DJ. Spindle dynamics during meiosis in Drosophila oocytes. J Cell Biol. 1997;137:1321–1336. [PMC free article: PMC2132525] [PubMed: 9182665]
- 51.
- Ashery-Padan R, Weiss AM, Feinstein N. et al. Distinct regions specify the targeting of otefin to the nucleoplasmic side of the nuclear envelope. J Biol Chem. 1997;272:2493–2499. [PubMed: 8999964]
- 52.
- Berman CL. The role of YA protein in Drosophila female meiosis 2000. MS Thesis, Cornell University: Ithaca, NY.
- 53.
- Nilson LA, Schupbach T. EGF receptor signaling in Drosophila oogenesis. Curr Top Dev Biol. 1999;44:203–243. [PubMed: 9891881]
- 54.
- Guillemin K, Williams T, Krasnow MA. A nuclear lamin is required for cytoplasmic organization and egg polarity in Drosophila. Nat Cell Biol. 2001;3:848–851. [PubMed: 11533666]
- 55.
- Fuller MT. Spermatogenesis, in The development of Drosophila melanogaster, Bate, M. and Martinez Arias, A., Editors. 1993. Cold Spring Harbor Laboratory Press: Plainview, New York. p.71–147.
- 56.
- Whalen AM, McConnell M, Fisher PA. Developmental regulation of Drosophila DNA topoisomerase II. J Cell Biol. 1991;112:203–213. [PMC free article: PMC2288815] [PubMed: 1846370]
- 57.
- Poccia D, Collas P. Transforming sperm nuclei into male pronuclei in vivo and in vitro, in Curr Top Dev Biol, Pederson, R.A. and Schatten, G., Editors. 1996. Academic Press: San Diego. p.25–88. [PubMed: 8787571]
- 58.
- Collas P, Poccia D. Remodeling the sperm nucleus into a male pronucleus at fertilization. Theriogenology. 1998;49:67–81. [PubMed: 10732122]
- 59.
- Perotti ME. Ultrastructural aspects of fertilization in Drosophila, in The functional anatomy of the spermatozoan, Proceedings of the Second International Symposium, Afzelins, Editor. 1975. Pergamon Press: Oxford. p.57–68.
- 60.
- Fitch KR, Wakimoto BT. The paternal effect gene ms(3)sneaky is required for sperm activation and the initiation of embryogenesis in Drosophila melanogaster. Dev Biol. 1998;197:270–282. [PubMed: 9630751]
- 61.
- Callaini G, Riparbelli MG. Fertilization in Drosophila melanogaster: centrosome inheritance and organization of the first mitotic spindle. Dev Biol. 1996;176:199–208. [PubMed: 8660861]
- 62.
- Crevel G, Cotterill S. DNA replication in cellfree extracts from Drosophila melanogaster. EMBO J. 1991;10:4361–4369. [PMC free article: PMC453189] [PubMed: 1756740]
- 63.
- Lopez JM, Wolfner MF. The developmentally regulated Drosophila embryonic nuclear lamina protein ‘Young Arrest’ (fs(1)Ya) is capable of associating with chromatin. J Cell Sci. 1997;110:643–651. [PubMed: 9092946]
- 64.
- Ulitzur N, Harel A, Goldberg M. et al. Nuclear membrane vesicle targeting to chromatin in a Drosophila embryo cellfree system. Mol Biol Cell. 1997;8:1439–1448. [PMC free article: PMC276168] [PubMed: 9285817]
- 65.
- Kawasaki K, Philpott A, Avilion AA. et al. Chromatin decondensation in Drosophila embryo extracts. J Biol Chem. 1994;269:10169–76. [PubMed: 8144519]
- 66.
- Crevel G, Huikeshoven H, Cotterill S. et al. Molecular and cellular characterization of CRP1, a Drosophila chromatin decondensation protein. J Struct Biol. 1997;118:9–22. [PubMed: 9087911]
- 67.
- Collas P, Poccia DL. Formation of the sea urchin male pronucleus in vitro: membrane-independent chromatin decondensation and nuclear envelope-dependent nuclear swelling. Mol Reprod Dev. 1995;42:106–113. [PubMed: 8562044]
- 68.
- Poccia D, Collas P. Nuclear envelope dynamics during male pronuclear development. Dev Growth Differ. 1997;39:541–550. [PubMed: 9338589]
- 69.
- Sonnenblick BD. The early embryology of Drosophila melanogaster, in The Biology of Drosophila, Demerec, M., Editor. 1950. John Wiley and Sons: New York. p.62–167.
- 70.
- O'Neill SL, Karr TL. Bidirectional incompatibility between conspecific populations of Drosophila simulans. Nature. 1990;348:178–180. [PubMed: 2234083]
- 71.
- Lassy CW, Karr TL. Cytological analysis of fertilization and early embryonic development in incompatible crosses of Drosophila simulans. Mech Dev. 1996;57:47–58. [PubMed: 8817452]
- 72.
- Callaini G, Dallai R, Riparbelli MG. Wolbachia-induced delay of paternal chromatin condensation does not prevent maternal chromosomes from entering anaphase in incompatible crosses of Drosophila simulans. J Cell Sci. 1997;110:271–80. [PubMed: 9044057]
- 73.
- Liu J, Song K, Wolfner MF. Mutational analyses of fs(1)Ya, an essential, developmentally regulated, nuclear envelope protein in Drosophila. Genetics. 1995;141:1473–1481. [PMC free article: PMC1206880] [PubMed: 8601487]
- 74.
- Lopez JL, Berman CL, Yu J. The YA mutation defines a control point between meiosis and mitosis in Drosophila. In preparation.
- 75.
- Yu J, Wolfner MF. Drosophila YA binds to DNA and histone H2B with four domains. Mol Biol Cell. 2002;13:558–569. [PMC free article: PMC65649] [PubMed: 11854412]
- 76.
- Goldberg M, Lu H, Stuurman N. et al. Interactions among Drosophila nuclear envelope proteins lamin, otefin, and YA. Mol Cell Biol. 1998;18:4315–23. [PMC free article: PMC109015] [PubMed: 9632815]
- 77.
- Tirian L, Timinszky G, Zhang C. Importin-beta is required for nuclear envelope assembly, submitted.
- 78.
- Timinszky G, Tirian L, Nagy FT. et al. The importin-beta P445L dominant negative mutant protein loses RanGTP binding ability and causes arrest during the exit from mitosis in Drosophila. J Cell Sci. 2002;115:1675–1687. [PubMed: 11950886]
- 79.
- Edgar BA, Datar SA. Zygotic degradation of two maternal Cdc25 mRNAs terminates Drosophila's early cell cycle program. Genes Dev. 1996;10:1966–1977. [PubMed: 8756353]
- 80.
- Edgar BA, Lehman DA, O'Farrell PH. Transcriptional regulation of string (cdc25): a link between developmental programming and the cell cycle. Development. 1994;120:3131–3143. [PMC free article: PMC2753456] [PubMed: 7720557]
- 81.
- Edgar BA, Sprenger F, Duronio RJ. et al. Distinct molecular mechanisms regulate cell cycle timing at successive stages of Drosophila embryogenesis. Genes Devel. 1994;8:440–452. [PMC free article: PMC6520052] [PubMed: 7510257]
- 82.
- Foe VE, Alberts BM. Studies of nuclear and cytoplasmic behaviour during the five mitotic cycles that precede gastrulation in Drosophila embryogenesis. J Cell Sci. 1983;61:31–70. [PubMed: 6411748]
- 83.
- Fogarty P, Campbell SD, Abu-Shumays R. et al. The Drosophila grapes gene is related to checkpoint gene chk1/rad27 and is required for late syncytial division fidelity. Curr Biol. 1997;7: 418–426. [PubMed: 9197245]
- 84.
- Yu KR, Saint RB, Sullivan W. The Grapes checkpoint coordinates nuclear envelope breakdown and chromosome condensation. Nat Cell Biol. 2000;2:609–615. [PubMed: 10980701]
- 85.
- Paddy MR, Saumweber H, Agard DA. et al. Time-resolved, in vivo studies of mitotic spindle formation and nuclear lamina breakdown in Drosophila early embryos. J Cell Sci. 1996;109:591–607. [PubMed: 8907705]
- 86.
- Steward R, Govind S. Dorsalventral polarity in the Drosophila embryo. Curr Opin Genet Dev. 1993;3:556–561. [PubMed: 8241766]
- 87.
- Lemosy EK, Hong CC, Hashimoto C. Signal transduction by a protease cascade. Trends Cell Biol. 1999;9:102–107. [PubMed: 10201075]
- 88.
- Jongens TA, Hay B, Jan LY. et al. The germcellless gene product: A posteriorly localized component necessary for germ cell development in Drosophila. Cell. 1992;70:569–584. [PubMed: 1380406]
- 89.
- Jongens TA, Ackerman LD, Swedlow JR. et al. Germcellless encodes a cell type-specific nuclear poreassociated protein and functions early in the germ cell specification pathway of Drosophila. Genes Dev. 1994;8:2123–2136. [PubMed: 7958883]
- 90.
- Nili E, Cojocaru GS, Kalma Y. et al. Nuclear membrane protein LAP2beta mediates transcriptional repression alone and together with its binding partner GCL (germcellless). J Cell Sci. 2001;114:3297–307. [PubMed: 11591818]
- Nuclear Envelope Dynamics in Drosophila Pronuclear Formation and in Embryos - Ma...Nuclear Envelope Dynamics in Drosophila Pronuclear Formation and in Embryos - Madame Curie Bioscience Database
- Cytokines in the Pathogenesis of Rheumatoid Arthritis and Collagen-Induced Arthr...Cytokines in the Pathogenesis of Rheumatoid Arthritis and Collagen-Induced Arthritis - Madame Curie Bioscience Database
- Evolutionary De Novo Design - Madame Curie Bioscience DatabaseEvolutionary De Novo Design - Madame Curie Bioscience Database
- The Evolution of CDK-Activating Kinases - Madame Curie Bioscience DatabaseThe Evolution of CDK-Activating Kinases - Madame Curie Bioscience Database
- Cell Cycle Reactivation in Skeletal Muscle and Other Terminally Differentiated C...Cell Cycle Reactivation in Skeletal Muscle and Other Terminally Differentiated Cells - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...