

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013.

X chromosome inactivation (XCI) is a process required to equalize the dosage of X-encoded genes between female and male cells. XCI is initiated very early during female embryonic development or upon differentiation of female embryonic stem (ES) cells and results in inactivation of one X chromosome in every female somatic cell. The regulation of XCI involves factors that also play a crucial role in ES cell maintenance and differentiation and the XCI process therefore provides a beautiful paradigm to study ES cell biology. In this chapter we describe the important cis and trans acting regulators of XCI and introduce the models that have been postulated to explain initiation of XCI in female cells only. We also discuss the proteins involved in the establishment of the inactive X chromosome and describe the different chromatin modifications associated with the inactivation process. Finally, we describe the potential of mouse and human ES and induced pluripotent stem (iPS) cells as model systems to study the XCI process.
Introduction
In many species, the sex of an individual is genetically determined by genes located on sex chromosomes.1 Mammals are heterogametic, in which the female nucleus contains two X chromosomes and a male nucleus one X chromosome and one Y chromosome. Sex chromosomes originated from a pair of autosomes and divergence of these autosomes in proto X and proto Y chromosomes was initiated by the emergence of the key male sex determining gene SRY, which evolved from the ancestral SOX3 gene on the proto Y chromosome.2-5 In subsequent steps the proto Y chromosome acquired genes involved in male fertility, resulting in a genomic region which was nonhomologous with the X chromosome. It is thought that the absence of homology initiated degeneration of the Y chromosome. Nowadays the Y chromosome contains only a limited number of genes, most of them involved in male fertility and sex determination.6,7 The X chromosome is still able to recombine in the female germ line, which prevented degradation and therefore resulted in the maintenance of a large chromosome containing more then a thousand genes, involved in a plethora of biological functions, varying from brain development to metabolism and fertility.1,8
Since both sexes contain an equal number of autosomal chromosomes, an equal dosage of X linked genes is needed to accomplish functional cell physiology. In placental mammals, dosage compensation of X linked genes between both sexes is achieved by inactivation of one of the two X chromosomes in females, in a process called X chromosome inactivation (XCI).9-11 XCI occurs during early female development and results in functional hetero-chromatinization and silencing of the X chromosome, which is maintained during subsequent cell divisions throughout life.12 Therefore in both sexes, only one X chromosome is functionally active. Expression of X encoded genes from this chromosome and the single X in male cells is twofold up regulated compared to autosomes, thereby further contributing to proper dosage compensation of X-encoded genes.13-16 In mouse, X chromosome inactivation (XCI) is present in two forms. In the extra-embryonic tissues XCI is imprinted, with the paternal X chromosome (Xp) being inactivated in all cells.17 This process is initiated very early during development, around the two- to eight-cell stage18,19 and is maintained in the developing extra-embryonic tissues of the embryo, including the fetal placenta. In contrast, the Xp is reactivated in the inner cell mass (ICM), which gives rise to the embryo proper, after which random X inactivation is initiated around day 5.5 of development (Fig. 1).
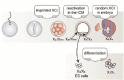
Embryonic stem (ES) cells are derived from the inner cell mass (ICM) of a blastocyst. They are characterized by the ability of self renewal and pluripotency with the capacity to form all cell types of the embryo proper and adult organism upon differentiation.20,21 Besides potential applications for regenerative medicine, ES cells are an ideal study system for early mammalian development from the pre-implantation period onwards.22,23 Female mouse ES cells retain two active X chromosomes (Xa) and upon differentiation these cells initiate random XCI, making them the prevailing model system to study XCI.24,25 Besides simulating early development, XCI in ES cells itself is important to accomplish proper cell function and developmental potential. The recent discovery of induced pluripotent stem (iPS) cells, which have ES cell characteristics and are derived from somatic cells by reprogramming with defined pluripotency factors,26-31 has reemphasized the importance of XCI. The active status of X chromosomes in ES and iPS cells can be used as a pluripotency marker, as during iPS reprogramming the inactivated X chromosome (Xi) from somatic cells becomes reactivated in iPS cells.32,33
In this chapter, we first describe the genetic key players involved in XCI, followed by a discussion of different models proposed to explain the counting and choice of X chromosomes during XCI and the establishment and maintenance of the inactive X during subsequent cell divisions. Finally, the differences between mouse and human ES cells regarding XCI are discussed.
Cis acting factors in XCI
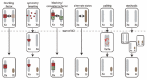
Transcriptional silencing of a whole chromosome during development has fascinated biologists for decades and in recent years a considerable amount of knowledge has been acquired contributing to our understanding of the molecular mechanisms involved in XCI. Genetic studies in mice and humans with X-to-autosome translocations have revealed that a major X-linked control locus, the X inactivation center (Xic in mice and XIC in humans) is necessary for XCI to occur.34-37 The Xic, which encompasses more then 1 Mb on the mouse X chromosome,38 has been shown to contain at least 4 genes which are involved in the process of XCI (Fig. 2). Three of these genes, Xist, Tsix and Xite, are noncoding and represent the master switch locus involved in silencing of the X chromosome in cis. The fourth gene, Rnf12 which will be discussed in later sections, encodes an E3 ubiquitin ligase involved in regulation of XCI in trans.

Figure 2
The X inactivation center on the X chromosome. Schematic representation of the mouse X chromosome and the location of part of the X inactivation center including the genes, Xist, Tsix, Xite and Rnf12 which are involved in regulating XCI.
The first gene which has been found to map to the XIC/Xic is the X-inactive specific transcript (Xist in mice, XIST in humans).37,39,40 Xist is the only known gene which is specifically expressed from the Xi. Xist is a noncoding gene, consisting of 7 exons in the mouse and 8 exons in humans, producing a poly-adenylated RNA molecule (17 kb in human and 15 kb in mouse), which is subject to alternative splicing.41,42 Xist RNA is tightly associated with the Xi43,44 and it is required for XCI to occur in cis, as knockout studies in female ES cells and mice have shown that X chromosomes bearing a deletion of the Xist gene are unable to inactivate the mutated X.45-47 During XCI, expression of Xist is up regulated from the future Xi.48,49 Xist RNA molecules spread from the Xic across the X chromosome in cis and cover the Xi completely. By spreading over the Xi, Xist RNA induces heterochromatinization of the X chromosome by attracting chromatin modifiers, involved in gene silencing.50-52
Another noncoding gene located within the Xic is Tsix, which is transcribed antisense to Xist.53 Tsix contains four exons and at least two transcriptional start sites, producing a 40 kb transcript, which only localizes to the Xic, as determined by RNA-FISH experiments. In mice the Tsix gene fully overlaps with Xist and Tsix has been shown to negatively regulate expression of Xist, as a deletion of Tsix leads to up regulation of Xist transcription and exclusive inactivation of the mutated X chromosome in female cells.54,55 Prior to XCI, expression of Tsix is from both X chromosomes in a 10 to 100 times excess compared to Xist and after initiation of XCI expression is continued transiently on the future Xa.56 On the Xi, transcriptional shutdown of Tsix is accompanied by chromatin changes at the Tsix promoter.57 The exact mechanisms involved in Tsix-mediated silencing of Xist are unknown. Since expression of Tsix transcripts is found in a gradient along the Xist gene, with more transcripts in the 5' portion of Tsix relative to the 3' portion of the gene, a role for transcriptional interference as a mechanism to suppress Xist has been proposed.49,56,58 Another possible mechanism by which Tsix might suppress Xist transcription is via RNA-mediated silencing. It has been shown that Tsix regulates the methylation status and thus the activity of the Xist promoter, via de novo methyltransferase 3a (DNMT3A).49 Also, active chromatin marks are more abundant at the Xist promoter in cells with a deficient Tsix gene in cis, whereas marks of repressed chromatin are reduced.59,60 Antisense transcription through the Xist promoter itself seems to be crucial for the establishment of repressive chromatin marks, as a truncation of Tsix to 93% of its normal length failed to induce Xist silencing.61 Also deleting the DXPas34 element, which is a CpG island located downstream to the major Tsix promoter and also initiates antisense transcription, abrogates Xist silencing in cis, thereby further emphasizing the importance of antisense transcription in Tsix-mediated silencing of Xist.60,62,63 Furthermore, the methylation status of this CpG island coincides perfectly with antisense transcription through Xist.64,65 Xist and Tsix transcripts are partially overlapping and therefore a possible role for an RNAi-mediated mechanism regulating XCI can not be excluded.66 Small xiRNAs, ranging in size from 25 to 42 nucleotides have indeed been detected from different regions within the Xist gene and a mutation of the endonuclease Dicer resulted in a loss of xiRNA formation and decreased methylation of Xist, implicating a role for Dicer in XCI. This is disputed by others, who found that Dicer null-ES cells show normal XCI and that the effects on the Xa are mediated by a decreased activity of de novo methyltransferases rather than a direct effect of Dicer.67,68 Therefore, at present the exact role of small RNAs in XCI initiation is unclear. Also, over expression in cis of Tsix cDNA in a cell line with abrogated endogenous Tsix transcription did not result in restoration of Tsix-mediated Xist silencing, which argues against an RNAi-mediated process.69
The third noncoding gene involved in XCI, Xite, for X chromosome intergenic transcript element,70 is located approximately 10 kb upstream of Tsix and its expression and methylation pattern during XCI is similar to that of Tsix. Xite is believed to be the positive regulator of Tsix.71 Deletion of Xite results in a reduced antisense transcription through the Xist locus, implying a similar role for Xite in inhibiting Xist expression as for Tsix and DXPas34.65,70
Trans acting factors in XCI
How are Xist, Tsix and Xite regulated? Among the proteins involved in Tsix regulation are the insulator protein CTCF and the transcription factor yin yang 1 (YY1), for which several tandemly organized binding sites have been identified in the DXpas34 region and the Xite promoter.72 Knockout studies involving Yy1, or partial ablation of Yy1 and Ctcf through RNAi mediated repression, revealed down-regulation of Tsix expression and concomitant up-regulation of Xist expression, supporting a role for YY1 and CTCF in Tsix expression.73 Recently, it has been shown that the pluripotency transcription factor OCT4 is able to bind to the DXpas34 element and the Xite promoter and may cooporate with CTCF in the regulation of Tsix transcription.74 Another study showed that the pluripotency factors NANOG, OCT4 and SOX275-78 can repress Xist expression by binding to a region within Xist intron 1.79 Nanog-deficient ES cells show up-regulation of Xist and this preceded down-regulation of Tsix, suggesting a Tsix-independent mechanism for the pluripotency factors in Xist repression. Interestingly, Nanog expression is also required for reactivation of the inactive paternal X chromosome in the ICM, most likely through suppression of Xist.80Therefore it seems likely that the well characterized key players of pluripotency are involved in repressing XCI prior to differentiation through different pathways.
A transgenesis screen in ES cells using BAC transgenes covering part of the Xic region, resulted in the identification of X-encoded Rnf12 as an activator of XCI.81 Rnf12 is located 500 kb telomeric from Xist, within the region that originally delineated the Xic and encodes an E3 ubiquitin ligase which has previously shown to be involved in the regulation of LIM-homeodomain transcription factors, estrogen dependent transcription activation and telomere length homeostasis.82-84 Additional copies of Rnf12 can induce ectopic XCI in male ES cells on the single X chromosome and in a significant proportion of female cells on both X chromosomes.81 Initiation of XCI was reduced in heterozygous Rnf12 female ES cells compared with wild type female ES cells. These findings indicate an important dose-dependent role for RNF12 in activation of XCI, although the molecular mechanism by which RNF12 mediates XCI remains elusive so far. Also, more unidentified activators of XCI must be present to explain initiation of XCI in female Rnf12 heterozygous knockout ES cells.
Counting and Choice
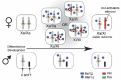
A long standing question in XCI research involved the counting and choice mechanism regulating initiation of XCI and required to inactivate the proper number of X chromosomes. How does a cell sense the number of X chromosomes present in a nucleus and how many of them need to be inactivated? A number of clinical observations in patients with an aberrant number of X chromosomes has shed some light on this question. In patients with a supernumerary number of X chromosomes, like so-called 46,XXX super females or 46, XXY Klinefelter patients, all but one X chromosome become inactivated.85-87 In Turner syndrome, the single X chromosome present in these females does not undergo XCI, whereas in tetraploid female embryos two inactivated X chromosomes are found.88,89 From this, the general rule has been deduced that XCI results in one Xa per diploid genome.90
Several models, with increasing complexity and partially overlapping mechanisms, have tried to explain these observations (Fig. 3). The blocking factor (BF) model predicts the presence of an autosomally encoded factor which is present in one entity in a diploid nucleus.35,36 The BF is thought to act through binding to a DNA element, called counting element and only interacts with one X chromosome, thereby rescuing this X from XCI in cis. Since there is only enough BF in a diploid nucleus to prevent XCI on one X chromosome, all additional X chromosomes will become silenced. The related symmetry breaking model states that the BF is not a single entity, but consists of several autosomally encoded molecules which assemble on the future Xa, thereby preventing XCI.91,92 Since the Xic is necessary for XCI to occur, as XCI is initiated only in the presence of more than one Xic in a diploid nucleus, it seems likely that the postulated counting element must be located within the Xic. Several studies made use of transgenes and deletions, to identify the counting element, reasoning that when an additional counting element would be introduced into male ES cells, this should be able to titrate away the limiting BF and induce counting. Therefore, XCI should also occur on the endogenous X chromosome, which is now no longer protected by the BF.
Indeed, several transgenic male ES cell lines have been generated with autosomally integrated transgenes covering Xist and Tsix, or Xist alone, in which ectopic XCI occurred on the single endogenous X chromosome.93-98 Autosomal Xist expression and silencing of autosomal genes adjacent to the integration site was also observed. The transgenes used varied from large YACs carrying >500 kb to small cosmids of only 35 kb covering only Xist and flanking regions.94 These studies therefore indicated that factors involved in counting may be located within the sequences covered by the transgenes. However, other related studies did not show induction of counting using similar transgenes,81,99,100 or showed that only multicopy transgenes are able to induce counting.101 Interestingly, studies involving a deletion of Xist on one X chromosome in female ES cells, which had shown that Xist is necessary for XCI to occur in cis, also showed that Xist transcription and the deleted part of the Xist gene are not involved in counting, as XCI is normally initiated on the wild type X chromosome.45-47,102-104 In contrast, different male Tsix mutant ES cells, some already generated prior to the identification of Tsix, displayed initiation of XCI on the single X chromosome, which suggested a role for the deleted sequences in the counting process.55,58,60,63,105 The same mutations did however not disturb the counting process in female cells, but resulted in preferential inactivation of the mutated allele, showing that Tsix mediates silencing of Xist in cis.54 These findings could be explained through a mechanism whereby the respective mutations disrupted the counting element, preventing BF binding. However, a heterozygous deletion of a region including Xist, Tsix and Xite (ΔXTX), in female ES cells and mice did not result in a disturbed counting process, as the wild type X chromosome was normally inactivated.106 This finding indicated that Xist, Tsix and Xite are not required for the counting process and locate the counting element outside the deleted region, although studies with Xist transgenes suggest that overlapping sequences may be involved which play a redundant role in counting and initiation of XCI.
In contrast to all previously reported Tsix mutations, one described Tsix mutation (ΔCpG), which involves a deletion of the DXPas34 region, did not result in aberrant XCI in male cells.107 Interestingly, female cells with a homozygous ΔCpG Tsix mutation showed chaotic XCI, with many cells initiating XCI on both X chromosomes. Based on this finding an X-encoded competence factor (CF) was hypothesized, involved in activation of XCI. One model comprising the combined action of a BF and CF postulates that the abundantly present CF inactivates all X chromosomes but not the one to which BF is bound.103 Another hypothesis states that also the X-encoded CF is limiting and is titrated away by one 'copy' of the autosomally-encoded BF, which corresponds to a single X chromosome.107 When more than one X chromosome is present in a diploid background, the extra copies of CF will not be titrated by the BF and will inactivate the remaining unprotected X chromosome(s)
The model of alternate states proposes that the two X chromosomes in an XX cell are already different prior to XCI.108 This model is supported by the fact that cohesion of sister chromatids has been shown to be differentially regulated between two X chromosomes in undifferentiated cells. Also differences in methylation and the chromatin state may play a role. Therefore an inherent epigenetic difference between two genetically identical chromosomes may exist prior to the initiation of XCI. However further experimental validations for these observation are needed.
A different model explains counting and choice in XCI by transient transvection or pairing events of the two Xic's present in a female diploid nucleus.109,110 This model is supported by observations that in early differentiating ES cells, there is a nonrandom spatial distribution of the Xic's in the nucleus, at which the Xic's move closer to each other prior to the onset of XCI. This transient pairing event may therefore play a role in the regulation of counting and choice. Pairing is facilitated by Tsix and Xite sequences and it seems to be dependent on the action of CTCF.111 The stem cell transcription factor Oct4 and transcription mediated by RNA polymerase II are also crucial for the pairing events.74,111 A genomic region containing part of the Slc16A2 gene, located 250 to 350 kb telomeric of Xist has been identified which also mediates pairing of the X chromosomes at the onset of XCI112 and was proposed to play a role in the activation of XCI. At present, it is unclear whether pairing has a functional role in XCI, or is a consequence of the transcriptional activation, which may result in relocation of the Xic in the nucleus. Interestingly, pairing appears not to be required for initiation of XCI, as XCI is initiated in XX-65kb ES cells with a deletion distal to Xist removing Tsix and Xite sequences, which abolish the XCI pairing event.109,110 Therefore additional studies are needed to clarify the role of pairing in XCI.
Many of the above discussed models assume that the XCI process is deterministic and mutually exclusive, in which always the correct number of X chromosomes are inactivated in female cells. However, in vitro studies with diploid and tetraploid ES and ICM cells revealed a significant percentage of cells with too many or too few Xi's,89,106,113 suggesting a stochastic mechanism directing the XCI process, with an independent probability for every X chromosome to initiate XCI.106 Comparison of the relative number of cells that initiated XCI between different diploid, triploid and tetraploid ES cells indicated that the X to autosome ratio determines the probability for an X chromosome to be inactivated.114 The probability is the resultant of different factors: X-encoded XCI-activators and autosomally encoded XCI-inhibitors that promote or repress Xist accumulation, respectively. Upon development or differentiation, the concentration of the XCI-activators will rise and/or the concentration of the XCI-inhibitors will decrease and in female cells this will be sufficient to generate a specific probability in time for enough Xist to accumulate and start to spread in cis (Fig. 4). XCI-inhibitors are involved in setting up a threshold that has to be overcome by Xist to accumulate. Because the XCI-activator gene is X-linked, spreading of Xist will down-regulate the XCI-activator gene in cis, preventing the second X chromosome from inactivation. In this model, initiation of spreading is a stochastic event, so that the chance for silencing of the XCI-activator gene on either X is equal. In male cells the concentration of the XCI-activator will not be sufficient to break the threshold and initiate XCI. Therefore, female specific initiation of XCI is obtained through a sex-dependent dosage difference in X-encoded XCI-activators that promote Xist accumulation. Cell lines and mice that harbor Xist or Tsix mutations that affect the expression of one of the genes indicate that Xist and Tsix are the major players in setting up the probability and that the XCI-activators and XCI-inhibitors are likely to act through these genes.
Several XCI-inhibitors have been identified, some of them acting through activation of Tsix expression, like Yy1, Ctcf and Oct4, or by direct suppression of Xist, like Nanog, Oct4 and Sox2.72-74,79 Rnf12 was identified as an X-encoded activator of XCI.81 The observed ectopic XCI induced by Rnf12 transgenes correlates with expression of transgenic Rnf12/RNF12, providing evidence for a dose-dependent role of RNF12 in activation of XCI. In agreement with this, heterozygous Rnf12 knockout female ES cells only initiate XCI in a significant reduced number of cells. RNF12 expression in wild type ES cells is up-regulated around the time XCI is initiated and prior to XCI, a twofold higher dosage of RNF12 protein has been detected in female compared to male ES cells. Rnf12 itself is subject to rapid silencing upon XCI induction, which will facilitate inhibition of initiation of XCI on the second X chromosome in female cells. How Rnf12 regulates XCI is unknown and future studies have to address the molecular mechanisms involved in Rnf12-mediated activation of XCI.
The recent identification of inhibitors and activators of XCI support a stochastic model for XCI. This indicates that the XCI process does not involve a choice mechanism, but is dictated by an initiation and feedback process, which is regulated through an intricate balance between inhibiting factors and activators of XCI.
Silencing and maintenance of silencing
Once XCI is initiated, a series of events takes place which changes the euchromatin of the active X chromosome into the tightly packed inactive heterochromatin of the Xi, which can be recognized as the Barr body in female somatic cells.115,116 The first step in this cascade of events is the transcriptional up regulation and spreading of Xist on the future Xi37,39 (Fig. 5). The Xist transcript contains several repeats, of which the A repeat, located in the 5'portion of Xist, is involved in the silencing process.117 Coating of the future Xi with Xist RNA leads to a rapid deprivation of RNA polymerase II and associated transcription factors, which leads to an immediate reduction of gene transcription on this chromosome and the creation of a silent nuclear compartment.118,119 One of the earliest events after Xist accumulation are specific chromatin changes, including the loss of active chromatin marks like H3K9 acetylation and H3K4 mono- and di-methylation,19,120,121 as well as a gain of silent chromatin marks like H3K27 tri-methylation (H3K27-me3),50,51,57,122 H3K9 di-methylation (H3K9-me2)121,123-126 and H4K20-mono-methylation (H4K20-me1),127 followed by the incorporation of histone variants including macroH2A128 and changes in DNA methylation and replication timing.129-131

To initiate the establishment of chromatin changes and silencing, Xist needs to accumulate and spread along the X chromosome. The exact mechanism involved in Xist spreading is not clear. Studies on X-to-autosome translocations and autosomally integrated Xist transgenes have indicated that Xist spreading on autosomal regions is not complete, which indicated that X-chromosomal sequences are needed for efficient spreading.35,95,132-135 Booster elements or way stations have been hypothesized to facilitate the Xist transmission along the X chromosome136 and LINE-1 retrotransposons were postulated to be the most likely candidate for being these booster elements,137-140 as they are one of the most common repetitive sequences in mammalian genomes and are enriched on the X chromosome compared to autosomes.141-144 Regions on the X chromosome that undergo XCI have a higher LINE-1 density than regions surrounding genes that escape inactivation8,142,143,145-147 and the density of LINE-1's on the fused autosome in X-to-autosome translocations seems to correlate with the amount of Xist spreading.133-135,148-153 However, also other sequences may be important for the Xist spreading, as others did not find a correlation between LINE-1 density and spreading, or suggested a less critical role.57,146,154-156 How Xist interacts with these sequences during the spreading process and whether these interactions are direct or mediated by other molecules is at present not clear.
After Xist accumulation, one of the first histone modifications detected on the Xi is H3K27-me3. This modification is dependent on the Polycomb repressive complex PRC2, which accumulates on the Xi and consists of the proteins EED, RbAp46/48, SUZ12 and EZH2, of which EZH2 is a methyl transferase implicated in H3K27 tri-methylation.51,157,158 Recently, a direct interaction between EZH2 and the A repeat of Xist has been reported and Ezh2 knockdown studies in female ES cells indicated a role for PRC2 in establishment of the Xi in random XCI.52 Interestingly, a knockout of EED resulting in a non functional PRC2 complex and the absence of H3K27-me3 only reveals a defect in imprinted XCI in mice,159 with re-activation of the Xi in extra-embryonic tissues. Despite defects in embryonic development due to the absence of EED, random XCI was not affected in the embryo,160 which contrasts with results obtained after knock down of Ezh2 in vitro.52 A different study, employing autosomally integrated inducible Xist transgenes, indicated that in the absence of EED some components of the PRC1 complex (MPH1 and MPH2) do not localize to the Xist coated autosome. However, recruitment of RING1B, which is part of the PRC1 complex and Xist mediated silencing of autosomal sequences were not affected.161 Accumulation of H2AK119ub1 (mono-ubiquitylation of histone H2A on lysine 119) on the Xi is dependent on RING1B,162,163 but also a homozygous Ring1b mutation does not affect Xist mediated silencing of an autosome with a transgenic insertion of Xist.161 These findings indicate that PRC1 and PRC2 are dispensable for random XCI, which does not exclude the possibility that these complexes are involved in redundant mechanisms in the establishment and maintenance of the Xi.
Also other proteins have been implicated in establishment and maintenance of the Xi. An E3 ubiquitin ligase complex consisting of the speckle-type POZ protein SPOP and CULLIN3 has been shown to be involved in the regulation of macroH2A deposition on the Xi.164 SPOP and CULLIN3 ubiquitinate both BMI1, a component of the PRC1 complex and macroH2A. Ubiquitination of macroH2A appears crucial for the recruitment of macroH2A to the Xi and RNAi-mediated knock-down of either SPOP or CULLIN3 results in diminished macroH2A staining on the Xi. RNAi-mediated knock-down of either macroH2A or SPOP/CULLIN3 in combination with demethylation and deacetylation inhibitor treatment, resulted in reactivation of an Xi-linked reporter gene. Reactivation was not found with a demethylation and deacetylation inhibitor treatment alone, indicating a role for macroH2A in the maintenance of the silent Xi state. The silencing function of macroH2A might be indirectly established by recruitment of the Poly (ADP-ribose) polymerase 1, PARP-1.165 PARP-1 is a nuclear enzyme involved in modulating chromatin structure, sensing DNA damage and regulation of gene expression.166-168 MacroH2A is able to recruit PARP-1 to the Xi and to inhibit the catalytic activity of PARP-1. Enzymatically inactive PARP-1 is able to bind nucleosomes and inhibit transcription.169 Depletion of PARP-1 in combination with demethylation and deacetylation inhibitor treatment leads to reactivation of an Xi-linked reporter gene. Hence, macroH2A might collaborate in gene silencing by modulating the enzymatic activity and thus the ability to silence, of PARP-1. The nuclear scaffold protein SAF-A is also enriched on the Xi. SAF-A has been characterized as a DNA/RNA binding protein and is a putative component of the nuclear scaffold involved in regulation of gene expression and DNA replication. Enrichment of SAF-A is dependent on the RNA binding domain of SAF-A, which might be involved in Xist mediated recruitment to the Xi.170 Recruitment of Xist to the Xi may be facilitated by BRCA1, a protein involved in many pathways including checkpoint activation and DNA repair. Association of BRCA1 with the Xi was found in a small percentage of cells and reconstitution experiments indicated a role for BRCA1 in Xist/XIST localization.171,172 Nonetheless, this claim is disputed by others and the exact role of BRCA1 in XCI remains to be determined.173,174 SMCHD1 also associates with the Xi and is involved in the methylation of CpG islands of genes subject to XCI.175 SMCHD1 contains a SMC-hinge domain which is found in proteins involved in cohesion and chromosome condensation. SmcHD1 knockout mice show defects in maintenance of the Xi in embryonic and extra-embryonic tissues. Analysis of Atrx heterozygous female knockout mice implicated a role for ATRX (a chromatin remodeling protein named alpha-thalassaemia and mental retardation on the X chromosome) in imprinted XCI.176 Female mice inheriting a mutated allele through the female germ line fail to inactivate the paternal X in extra-embryonic tissues, suggesting a failure in imprinted XCI. Interestingly, at later stages of ES cell differentiation, ATRX also associates with the Xi, supporting a role in maintenance of the Xi.177 SatB1 and SatB2 have also been implicated in the establishment of the Xi. These cancer associated genes encode nuclear proteins, which act as genome organizers and gene regulators.178 Knockdown studies of SatB1 and SatB2 showed a partial defect in silencing of the X chromosome in differentiating female ES cells, suggesting a direct role for these proteins in heterochromatinization of the Xi.179
Interestingly, once the Xi is established, expression of Xist is no longer necessary for the maintenance, as a conditional deletion of Xist does not lead to reactivation of silenced genes.47 Multiple redundant proteins and epigenetic layers are thus involved in the establishment and maintenance of the Xi. Together, they ensure that the Xi is stably silenced, leading to persistent dosage compensation. Only a limited number of X-linked genes is able to escape XCI mediated silencing in the mouse. In humans, more escapees have been identified.180-183 Therefore, in different species, the robustness of silencing might differ, which might be caused by a differential evolutionary need for dosage compensation between species. The genes which escape dosage compensation and do not have Y-chromosomal homologues, are thus differentially expressed between males and females and may play a role in establishing differences between both sexes.
In summary, many different redundant epigenetic layers and only partly understood complex interactions are involved in the silencing and maintenance of the silent state of the X chromosome. Although many investigations have focussed on these mechanisms, only the first steps have been made towards a complete understanding of the silencing and maintenance of XCI.
XCI and human ES cells
Studying X chromosome inactivation in humans is challenging. Due to ethical reasons, the use of early human embryos for research purposes has been widely restricted. Therefore, most of the obtained knowledge on human XCI has come from studies which made use of different model systems, including mouse-human cell hybrids, human embryonal carcinoma and tumour cell lines184,185 and human transgenes integrated in mouse ES cells.81,97,186-188 The derivation of human ES (hES) cells promised the availability of a potent study model for human XCI, comparable to mouse ES cells. Different studies explored XCI in hES cells, with varying and conflicting results.189-194 Most of the female hES cell lines display an inactivated X chromosome already in the undifferentiated state characterized by XIST expression, XIST coating and accumulated markers of heterochromatin on the Xi. Other undifferentiated cell lines have active X chromosomes and have the potential to inactivate during differentiation, comparable to mouse ES cells (Table 1). Interestingly, certain cell lines behave different in distinct laboratories, with some sub clones showing random XCI upon differentiation, whereas others show XCI hallmarks prior to differentiation. In a survey of 11 characterized hES cell lines, Silva et al identified three different classes of hES cells with regard to XCI.191 The first class only displays XCI characteristics upon differentiation. The second and third classes have an inactive X chromosome in the undifferentiated state, but in the third group XIST expression is lost. Although this last category does no longer express XIST, other XCI markers are still present, like the exclusion of Cot-1 RNA from the X chromosome. Interestingly, in these cells H3K27 tri-methylation is also lost, which indicates that H3K27 tri-methylation is dispensable for the maintenance of XCI. These results support previous findings that in mice and human recruitment of H3K27 tri-methylation is XIST dependent.50
Table 1
Characteristics of mouse and human pluripotent stem cells.
How can the differences between hES and mouse ES cells regarding XCI be explained? In mice, cells of the ICM show two Xa's prior to differentiation. A species specific difference in the time window in which XCI occurs could explain why hES cells, which are also derived from the ICM, display XCI characteristics prior to differentiation. Does random XCI in human embryos already occur at an earlier stage and do hES cells therefore display an Xi? Alternatively, reactivation of the Xi established during the early cleavage stages might occur later in humans compared to mouse. In mice, imprinted XCI occurs prior to implantation and causes inactivation of the Xp.17-19,195,196 Imprinted XCI is maintained in the extra-embryonic tissues, but is reversed in the cells of the ICM (Fig. 1), by reactivating the Xp, most likely initiated by Nanog expression in cells of the epiblast.80 Although in human XCI is also initiated during the early cleavage divisions,197 it is unclear whether human XCI is imprinted.198-207 In addition it is unclear at what stage the Xi is reactivated, if reactivated at all and hence, XCI in undifferentiated hES cells could be due to persistence of XCI from the early pre-implantation embryo, rather then precocious initiation of random XCI. In support of this, a highly skewed XCI pattern has been observed in female hES cells with a preference for one of the two X chromosomes to be inactivated.190,208 Unfortunately, the paternal origin of the inactivated X chromosome was not determined.
Beside variations in the XCI timing between species, the observed differences in XCI between undifferentiated human and mouse ES cells could also be explained by fundamental differences between the two cell types.209,210 Human and mouse ES cells differ significantly in morphology, clonogenicity, molecular profile and culture requirements. For example, hES cells need bFGF and Activin/Nodal signalling for their self renewal, whereas mouse ES cells depend on LIF/Stat3 signalling211,212 and both cell types are characterized by a genome wide difference in pluripotency factor promoter occupancy.213 Although both cell types have the potential to differentiate in vitro into cell types of all germ layers and to form teratomas in vivo, only for mouse ES cells the capability to contribute to the germ line and generate an entire animal has been tested. For hES cells the generation of chimaeras is not possible due to ethical reasons. Therefore at present it is unclear whether hES cells are indeed the proper human equivalent of mouse ES cells. In mice, an additional pluripotent cell population has been isolated from the post-implantation mouse epiblast using culture conditions including bFGF and Activin, in the absence of LIF.214,215 These so-called EpiSCs (post-implantation epiblast derived stem cells) express the pluripotency factors Oct4, Sox2 and Nanog, but differ in morphology and in the expression of certain transcripts from mouse ES cells, making them more comparable to hES cells.216 Although these cells can differentiate in vitro into cells of all germ layers and extra-embryonic trophectoderm, they are extremely inefficient in contributing to chimaeras and germ line transmission has not been reported. Interestingly, female EpiSCs have undergone XCI, leaving one X chromosome active. Therefore, it is possible that XCI observed in hES cells is a reflection of the difference between human and mouse ES cells, with hES cells being in fact the human counterpart of mouse EpiSCs.217 In agreement with this, mouse EpiSCs and hES cells share the ability to differentiate in vitro into extra-embryonic tissues,218 whereas mouse ES cells can only differentiate into cells of the three germ layers. It has been shown that EpiSCs can be differentiated from mouse ES cells in culture, which indicates that EpiSCs are a more restricted derivative from ES cells.219 Also reprogramming of EpiSCs to ES cells by over expression of the pluripotency factor Klf4,219 Nanog80 or extended culture in LIF,220 is accompanied by the reactivation of the inactive X chromosome. How can human ES cells be the human equivalent of a more differentiated cell type, despite the fact that human ES cells are also derived from the ICM, like mouse ES cells? It is possible that the culture conditions used for the derivation of hES cells allow the differentiation of ground state hES cells to hES cells with EpiSCs characteristics and prevent in vitro stabilization of the pluripotent state of the human ICM.221 Also differences in early development between rodents and humans, such as the presence of an egg cylinder or the presence of a temporary arrest in embryonic development called diapause specific for the mouse, might explain why in humans a shorter time window may exist to derive ground state ES cells, comparable to mice.222 Therefore it is possible that during human ES cell derivation in fact only more differentiated cells are derived, which might therefore explain XCI in these cells.
The presence of an inactive X in undifferentiated hES cells could also be explained by a selective pressure against two active X chromosomes. In fact, during development, only a short time window exists in which cells in the early embryo have two active X chromosomes. Since these cells are not programmed for an infinitive state in the ICM, these cells might adapt to culture conditions during ES cell derivation by initiating XCI, which might confer them with a growth or survival advantage. Selection against two active X chromosomes is also seen in some mouse ES cell lines and female mouse ES cells show genome wide hypomethylation,223 possibly resulting in genome instability. This may explain why many inbred mouse ES cells loose one of the two X chromosomes during expansion and only ES cells from hybrid crosses stably propagate two X chromosomes. Since hES cells are not characterized by a tendency to X chromosomal loss, they might employ XCI to prevent activity of two X chromosomes.
Taken together, the finding that XCI is already initiated in undifferentiated hES cells could have many different reasons and most likely a combination of these mechanisms will turn out to explain the epigenetic fluidity observed in hES cells. Future studies are needed to determine the origin of this finding, since concerns about the epigenetic stability of hES cells and differentiation potential of these cells, should be clarified before the introduction of hES cells for any clinical application.191,224-226
Could the recent emergence of human iPS cells provide us with a better model to study XCI? For mouse iPS cells, reactivation of the silent X chromosome of somatic cells during reprogramming has been shown32,33 and these cells initiate XCI upon differentiation. Although until today no studies have addressed the XCI status in human iPS cells, preliminary results from our laboratory indicate that also in human iPS cells precocious XIST accumulation is observed (T.S.B and J.G unpublished observations). Whether this is caused by culture artefacts, fundamental differences in the pluripotent state of human ES and iPS cells compared to mouse, or failure of complete reprogramming needs to be clarified in future investigations.
Conclusion
Mouse ES cells provide a powerful model system to study XCI, as XCI is initiated in female ES cells upon differentiation. Recent findings indicate that XCI is suppressed in female ES through the action of Nanog, Sox2 and Oct4, which also play a key role in maintaining the pluripotent ground state of ES cells as part of a complex transcription factor network. Activation of XCI requires down regulation of these factors and up-regulation of X-encoded activators of XCI, including Rnf12 which is also involved in differentiation processes. This indicates that XCI is regulated through the same factors that are also involved in pluripotency and differentiation. Nevertheless, many factors regulating XCI and the downstream targets of these factors still need to be identified. The tight link between pluripotency and XCI underscores that the epigenetic status of the X chromosome provides a potent readout to study pluripotency and nuclear reprogramming of female mouse cells. Unfortunately, for human female ES cells the picture appears to be more complex and it remains unclear whether female human ES cells provide a true model system to study the XCI process. Clearly more studies have to be performed to solve this issue.
Acknowledgements
We would like to thank all members of the laboratory for helpful and stimulating discussions. This work was supported by a NWO TOP grant to J.G. and a grant from the Dutch government (BSIK programme 03038, SCDD).
References
- 1.
- Ohno S, editor. Berlin: Springer; 1967. Sex chromosomes and Sex-linked genes.
- 2.
- Charlesworth B. The evolution of sex chromosomes. Science. 1991;251(4997):1030–3. [PubMed: 1998119]
- 3.
- Graves JA. The origin and function of the mammalian Y chromosome and Y-borne genes—an evolving understanding. Bioessays. 1995;17(4):311–20. [PubMed: 7741724]
- 4.
- Graves JA, Koina E, Sankovic N. How the gene content of human sex chromosomes evolved. Curr Opin Genet Dev. 2006;16(3):219–24. [PubMed: 16650758]
- 5.
- Graves JA. Sex chromosome specialization and degeneration in mammals. Cell. 2006;124(5):901–14. [PubMed: 16530039]
- 6.
- Skaletsky H, et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature. 2003;423(6942):825–37. [PubMed: 12815422]
- 7.
- Koopman P, et al. Male development of chromosomally female mice transgenic for Sry. Nature. 1991;351(6322):117–21. [PubMed: 2030730]
- 8.
- Ross MT, et al. The DNA sequence of the human X chromosome. Nature. 2005;434(7031):325–37. [PMC free article: PMC2665286] [PubMed: 15772651]
- 9.
- Lyon MF. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature. 1961;190:372–3. [PubMed: 13764598]
- 10.
- Lyon MF, Phillips RJ, Searle AG. A test for mutagenicity of caffeine in mice. Z Vererbungsl. 1962;93:7–13. [PubMed: 14467628]
- 11.
- Lyon MF. Sex chromatin and gene action in the mammalian X-chromosome. Am J Hum Genet. 1962;14:135–48. [PMC free article: PMC1932279] [PubMed: 14467629]
- 12.
- Davidson RG, Nitowsky HM, Childs B. Demonstration of two populations of cells in the human female heterozygous for glucose-6-phosphate dehydrogenase variants. Proc Natl Acad Sci USA. 1963;50:481–5. [PMC free article: PMC221205] [PubMed: 14067093]
- 13.
- Adler DA, et al. Evidence of evolutionary up-regulation of the single active X chromosome in mammals based on Clc4 expression levels in Mus spretus and Mus musculus. Proc Natl Acad Sci USA. 1997;94(17):9244–8. [PMC free article: PMC23138] [PubMed: 9256467]
- 14.
- Birchler JA, Fernandez HR, Kavi HH. Commonalities in compensation. Bioessays. 2006;28(6):565–8. [PubMed: 16700062]
- 15.
- Lin H, et al. Dosage compensation in the mouse balances up-regulation and silencing of X-linked genes. PLoS Biol. 2007;5(12):e326. [PMC free article: PMC2121114] [PubMed: 18076287]
- 16.
- Nguyen DK, Disteche CM. Dosage compensation of the active X chromosome in mammals. Nat Genet. 2006;38(1):47–53. [PubMed: 16341221]
- 17.
- Takagi N, Sasaki M. Preferential inactivation of the paternally derived X chromosome in the extraembryonic membranes of the mouse. Nature. 1975;256(5519):640–2. [PubMed: 1152998]
- 18.
- Huynh KD, Lee JT. Inheritance of a pre-inactivated paternal X chromosome in early mouse embryos. Nature. 2003;426(6968):857–62. [PubMed: 14661031]
- 19.
- Okamoto I, et al. Epigenetic dynamics of imprinted X inactivation during early mouse development. Science. 2004;303(5658):644–9. [PubMed: 14671313]
- 20.
- Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292(5819):154–6. [PubMed: 7242681]
- 21.
- Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci USA. 1981;78(12):7634–8. [PMC free article: PMC349323] [PubMed: 6950406]
- 22.
- Thomson JA, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282(5391):1145–7. [PubMed: 9804556]
- 23.
- Keller G. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev. 2005;19(10):1129–55. [PubMed: 15905405]
- 24.
- Leahy A, et al. Use of developmental marker genes to define temporal and spatial patterns of differentiation during embryoid body formation. J Exp Zool. 1999;284(1):67–81. [PubMed: 10368935]
- 25.
- Monk M. A stem-line model for cellular and chromosomal differentiation in early mouse-development. Differentiation. 1981;19(2):71–6. [PubMed: 7274618]
- 26.
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663–76. [PubMed: 16904174]
- 27.
- Nakagawa M, et al. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat Biotechnol. 2008;26(1):101–6. [PubMed: 18059259]
- 28.
- Wernig M, et al. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature. 2007;448(7151):318–24. [PubMed: 17554336]
- 29.
- Yu J, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917–20. [PubMed: 18029452]
- 30.
- Takahashi K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–72. [PubMed: 18035408]
- 31.
- Park IH, et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008;451(7175):141–6. [PubMed: 18157115]
- 32.
- Maherali N, et al. Directly reprogrammed fibroblasts show global epigenetic remodeling and widespread tissue contribution. Cell Stem Cell. 2007;1(1):55–70. [PubMed: 18371336]
- 33.
- Stadtfeld M, et al. Defining molecular cornerstones during fibroblast to iPS cell reprogramming in mouse. Cell Stem Cell. 2008;2(3):230–40. [PMC free article: PMC3538379] [PubMed: 18371448]
- 34.
- Russell LB. Mammalian X-chromosome action: inactivation limited in spread and region of origin. Science. 1963;140:976–8. [PubMed: 13975649]
- 35.
- Rastan S. Non-random X-chromosome inactivation in mouse X-autosome translocation embryos—location of the inactivation centre. J Embryol Exp Morphol. 1983;78:1–22. [PubMed: 6198418]
- 36.
- Rastan S, Robertson EJ. X-chromosome deletions in embryo-derived (EK) cell lines associated with lack of X-chromosome inactivation. J Embryol Exp Morphol. 1985;90:379–88. [PubMed: 3834036]
- 37.
- Brown CJ, et al. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature. 1991;349(6304):38–44. [PubMed: 1985261]
- 38.
- Heard E, Avner P. Role play in X-inactivation. Hum Mol Genet. 1994 Mar;Spec No:1481–5. [PubMed: 7849742]
- 39.
- Borsani G, et al. Characterization of a murine gene expressed from the inactive X chromosome. Nature. 1991;351(6324):325–9. [PubMed: 2034278]
- 40.
- Brockdorff N, et al. Conservation of position and exclusive expression of mouse Xist from the inactive X chromosome. Nature. 1991;351(6324):329–31. [PubMed: 2034279]
- 41.
- Brockdorff N, et al. The product of the mouse Xist gene is a 15 kb inactive X-specific transcript containing no conserved ORF and located in the nucleus. Cell. 1992;71(3):515–26. [PubMed: 1423610]
- 42.
- Brown CJ, et al. The human XIST gene: analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell. 1992;71(3):527–42. [PubMed: 1423611]
- 43.
- Clemson CM, et al. XIST RNA paints the inactive X chromosome at interphase: evidence for a novel RNA involved in nuclear/chromosome structure. J Cell Biol. 1996;132(3):259–75. [PMC free article: PMC2120729] [PubMed: 8636206]
- 44.
- Jonkers I, et al. Xist RNA is confined to the nuclear territory of the silenced X chromosome throughout the cell cycle. Mol Cell Biol. 2008;28(18):5583–94. [PMC free article: PMC2546918] [PubMed: 18625719]
- 45.
- Penny GD, et al. Requirement for Xist in X chromosome inactivation. Nature. 1996;379(6561):131–7. [PubMed: 8538762]
- 46.
- Marahrens Y, et al. Xist-deficient mice are defective in dosage compensation but not spermatogenesis. Genes Dev. 1997;11(2):156–66. [PubMed: 9009199]
- 47.
- Csankovszki G, et al. Conditional deletion of Xist disrupts histone macroH2A localization but not maintenance of X inactivation. Nat Genet. 1999;22(4):323–4. [PubMed: 10431231]
- 48.
- Kay GF, et al. Expression of Xist during mouse development suggests a role in the initiation of X chromosome inactivation. Cell. 1993;72(2):171–82. [PubMed: 8425217]
- 49.
- Sun BK, Deaton AM, Lee JT. A transient heterochromatic state in Xist preempts X inactivation choice without RNA stabilization. Mol Cell. 2006;21(5):617–28. [PubMed: 16507360]
- 50.
- Plath K, et al. Role of histone H3 lysine 27 methylation in X inactivation. Science. 2003;300(5616):131–5. [PubMed: 12649488]
- 51.
- Silva J, et al. Establishment of histone h3 methylation on the inactive X chromosome requires transient recruitment of Eed-Enx1 polycomb group complexes. Dev Cell. 2003;4(4):481–95. [PubMed: 12689588]
- 52.
- Zhao J, et al. Polycomb proteins targeted by a short repeat RNA to the mouse X chromosome. Science. 2008;322(5902):750–6. [PMC free article: PMC2748911] [PubMed: 18974356]
- 53.
- Lee JT, Davidow LS, Warshawsky D. Tsix, a gene antisense to Xist at the X-inactivation centre. Nat Genet. 1999;21(4):400–4. [PubMed: 10192391]
- 54.
- Lee JT, Lu N. Targeted mutagenesis of Tsix leads to nonrandom X inactivation. Cell. 1999;99(1):47–57. [PubMed: 10520993]
- 55.
- Clerc P, Avner P. Role of the region 3 to Xist exon 6 in the counting process of X-chromosome inactivation. Nat Genet. 1998;19(3):249–53. [PubMed: 9662396]
- 56.
- Shibata S, Lee JT. Characterization and quantitation of differential Tsix transcripts: implications for Tsix function. Hum Mol Genet. 2003;12(2):125–36. [PubMed: 12499393]
- 57.
- Marks H, et al. High-resolution analysis of epigenetic changes associated with X inactivation. Genome Res. 2009;19(8):1361–73. [PMC free article: PMC2720191] [PubMed: 19581487]
- 58.
- Luikenhuis S, Wutzand A, Jaenisch R. Antisense transcription through the Xist locus mediates Tsix function in embryonic stem cells. Mol Cell Biol. 2001;21(24):8512–20. [PMC free article: PMC100014] [PubMed: 11713286]
- 59.
- Navarro P, et al. Tsix transcription across the Xist gene alters chromatin conformation without affecting Xist transcription: implications for X-chromosome inactivation. Genes Dev. 2005;19(12):1474–84. [PMC free article: PMC1151664] [PubMed: 15964997]
- 60.
- Vigneau S, et al. An essential role for the DXPas34 tandem repeat and Tsix transcription in the counting process of X chromosome inactivation. Proc Natl Acad Sci USA. 2006;103(19):7390–5. [PMC free article: PMC1464350] [PubMed: 16648248]
- 61.
- Ohhata T, et al. Crucial role of antisense transcription across the Xist promoter in Tsix-mediated Xist chromatin modification. Development. 2008;135(2):227–35. [PubMed: 18057104]
- 62.
- Cohen DE, et al. The DXPas34 repeat regulates random and imprinted X inactivation. Dev Cell. 2007;12(1):57–71. [PubMed: 17199041]
- 63.
- Debrand E, et al. Functional analysis of the DXPas34 locus, a 3 regulator of Xist expression. Mol Cell Biol. 1999;19(12):8513–25. [PMC free article: PMC84967] [PubMed: 10567576]
- 64.
- Prissette M, et al. Methylation profiles of DXPas34 during the onset of X-inactivation. Hum Mol Genet. 2001;10(1):31–8. [PubMed: 11136711]
- 65.
- Boumil RM, et al. Differential methylation of Xite and CTCF sites in Tsix mirrors the pattern of X-inactivation choice in mice. Mol Cell Biol. 2006;26(6):2109–17. [PMC free article: PMC1430288] [PubMed: 16507990]
- 66.
- Ogawa Y, Sun BK, Lee JT. Intersection of the RNA interference and X-inactivation pathways. Science. 2008;320(5881):1336–41. [PMC free article: PMC2584363] [PubMed: 18535243]
- 67.
- Nesterova TB, et al. Dicer regulates Xist promoter methylation in ES cells indirectly through transcriptional control of Dnmt3a. Epigenetics Chromatin. 2008;1(1):2. [PMC free article: PMC2577046] [PubMed: 19014663]
- 68.
- Kanellopoulou C, et al. X chromosome inactivation in the absence of Dicer. Proc Natl Acad Sci USA. 2009;106(4):1122–7. [PMC free article: PMC2633553] [PubMed: 19164542]
- 69.
- Shibata S, Lee JT. Tsix transcription- versus RNA-based mechanisms in Xist repression and epigenetic choice. Curr Biol. 2004;14(19):1747–54. [PubMed: 15458646]
- 70.
- Ogawa Y, Lee JT. Xite, X-inactivation intergenic transcription elements that regulate the probability of choice. Mol Cell. 2003;11(3):731–43. [PubMed: 12667455]
- 71.
- Stavropoulos N, Rowntree RK, Lee JT. Identification of developmentally specific enhancers for Tsix in the regulation of X chromosome inactivation. Mol Cell Biol. 2005;25(7):2757–69. [PMC free article: PMC1061649] [PubMed: 15767680]
- 72.
- Chao W, et al. CTCF, a candidate trans-acting factor for X-inactivation choice. Science. 2002;295(5553):345–7. [PubMed: 11743158]
- 73.
- Donohoe ME, et al. Identification of a Ctcf cofactor, Yy1, for the X chromosome binary switch. Mol Cell. 2007;25(1):43–56. [PubMed: 17218270]
- 74.
- Donohoe ME, et al. The pluripotency factor Oct4 interacts with Ctcf and also controls X-chromosome pairing and counting. Nature. 2009;460(7251):128–32. [PMC free article: PMC3057664] [PubMed: 19536159]
- 75.
- Chambers I, et al. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003;113(5):643–55. [PubMed: 12787505]
- 76.
- Mitsui K, et al. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell. 2003;113(5):631–42. [PubMed: 12787504]
- 77.
- Nichols J, et al. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell. 1998;95(3):379–91. [PubMed: 9814708]
- 78.
- Avilion AA, et al. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 2003;17(1):126–40. [PMC free article: PMC195970] [PubMed: 12514105]
- 79.
- Navarro P, et al. Molecular coupling of Xist regulation and pluripotency. Science. 2008;321(5896):1693–5. [PubMed: 18802003]
- 80.
- Silva J, et al. Nanog is the gateway to the pluripotent ground state. Cell. 2009;138(4):722–37. [PMC free article: PMC3437554] [PubMed: 19703398]
- 81.
- Jonkers I, et al. RNF12 is an X-Encoded dose-dependent activator of X chromosome inactivation. Cell. 2009;139(5):999–1011. [PubMed: 19945382]
- 82.
- Bach I, et al. RLIM inhibits functional activity of LIM homeodomain transcription factors via recruitment of the histone deacetylase complex. Nat Genet. 1999;22(4):394–9. [PubMed: 10431247]
- 83.
- Her YR, Chung IK. Ubiquitin Ligase RLIM Modulates Telomere Length Homeostasis through a Proteolysis of TRF1. J Biol Chem. 2009;284(13):8557–66. [PMC free article: PMC2659214] [PubMed: 19164295]
- 84.
- Johnsen SA, et al. Regulation of estrogen-dependent transcription by the LIM cofactors CLIM and RLIM in breast cancer. Cancer Res. 2009;69(1):128–36. [PMC free article: PMC2713826] [PubMed: 19117995]
- 85.
- Jacobs PA, et al. Evidence for the existence of the human "super female". Lancet. 1959;2(7100):423–5. [PubMed: 14406377]
- 86.
- Maclean N, Mitchell JM. A survey of sex-chromosome abnormalities among 4514 mental defectives. Lancet. 1962;1(7224):293–6. [PubMed: 14468012]
- 87.
- Grumbach MM, Morishima A, Taylor JH. Human Sex Chromosome Abnormalities in Relation to DNA Replication and Heterochromatinization. Proc Natl Acad Sci USA. 1963;49(5):581–9. [PMC free article: PMC299924] [PubMed: 16591069]
- 88.
- Carr DH. Chromosome studies in selected spontaneous abortions. 1. Conception after oral contraceptives. Can Med Assoc J. 1970;103(4):343–8. [PMC free article: PMC1930439] [PubMed: 5447713]
- 89.
- Webb S, de Vries TJ, Kaufman MH. The differential staining pattern of the X chromosome in the embryonic and extraembryonic tissues of postimplantation homozygous tetraploid mouse embryos. Genet Res. 1992;59(3):205–14. [PubMed: 1380938]
- 90.
- Harnden DG. Nuclear sex in triploid XXY human cells. Lancet. 1961;2(7200):488. [PubMed: 13711655]
- 91.
- Nicodemi M, Prisco A. Symmetry-breaking model for X-chromosome inactivation. Phys Rev Lett. 2007;98(10):108104. [PubMed: 17358571]
- 92.
- Nicodemi M, Prisco A. Self-assembly and DNA binding of the blocking factor in x chromosome inactivation. PLoS Comput Biol. 2007;3(11):e210. [PMC free article: PMC2065890] [PubMed: 17997592]
- 93.
- Lee JT, et al. A 450 kb transgene displays properties of the mammalian X-inactivation center. Cell. 1996;86(1):83–94. [PubMed: 8689690]
- 94.
- Herzing LB, et al. Xist has properties of the X-chromosome inactivation centre. Nature. 1997;386(6622):272–5. [PubMed: 9069284]
- 95.
- Lee JT, Jaenisch R. Long-range cis effects of ectopic X-inactivation centres on a mouse autosome. Nature. 1997;386(6622):275–9. [PubMed: 9069285]
- 96.
- Lee JT, Lu N, Han Y. Genetic analysis of the mouse X inactivation center defines an 80-kb multifunction domain. Proc Natl Acad Sci USA. 1999;96(7):3836–41. [PMC free article: PMC22381] [PubMed: 10097124]
- 97.
- Migeon BR, et al. Human X inactivation center induces random X chromosome inactivation in male transgenic mice. Genomics. 1999;59(2):113–21. [PubMed: 10409422]
- 98.
- Migeon BR, et al. Low-copy-number human transgene is recognized as an X inactivation center in mouse ES cells, but fails to induce cis-inactivation in chimeric mice. Genomics. 2001;71(2):156–62. [PubMed: 11161809]
- 99.
- Heard E, et al. Transgenic mice carrying an Xist-containing YAC. Hum Mol Genet. 1996;5(4):441–50. [PubMed: 8845836]
- 100.
- Matsuura S, et al. Xist expression from an Xist YAC transgene carried on the mouse Y chromosome. Hum Mol Genet. 1996;5(4):451–9. [PubMed: 8845837]
- 101.
- Heard E, et al. Xist yeast artificial chromosome transgenes function as X-inactivation centers only in multicopy arrays and not as single copies. Mol Cell Biol. 1999;19(4):3156–66. [PMC free article: PMC84109] [PubMed: 10082582]
- 102.
- Gribnau J, et al. X chromosome choice occurs independently of asynchronous replication timing. J Cell Biol. 2005;168(3):365–73. [PMC free article: PMC2171734] [PubMed: 15668296]
- 103.
- Marahrens Y, Loring J, Jaenisch R. Role of the Xist gene in X chromosome choosing. Cell. 1998;92(5):657–64. [PubMed: 9506520]
- 104.
- Nesterova TB, et al. Skewing X chromosome choice by modulating sense transcription across the Xist locus. Genes Dev. 2003;17(17):2177–90. [PMC free article: PMC196458] [PubMed: 12952890]
- 105.
- Sado T, Li E, Sasaki H. Effect of TSIX disruption on XIST expression in male ES cells. Cytogenet Genome Res. 2002;99(1-4):115–8. [PubMed: 12900553]
- 106.
- Monkhorst K, et al. X inactivation counting and choice is a stochastic process: evidence for involvement of an X-linked activator. Cell. 2008;132(3):410–21. [PubMed: 18267073]
- 107.
- Lee JT. Homozygous Tsix mutant mice reveal a sex-ratio distortion and revert to random X-inactivation. Nat Genet. 2002;32(1):195–200. [PubMed: 12145659]
- 108.
- Mlynarczyk-Evans S, et al. X chromosomes alternate between two states prior to random X-inactivation. PLoS Biol. 2006;4(6):e159. [PMC free article: PMC1457015] [PubMed: 16669701]
- 109.
- Bacher CP, et al. Transient colocalization of X-inactivation centres accompanies the initiation of X inactivation. Nat Cell Biol. 2006;8(3):293–9. [PubMed: 16434960]
- 110.
- Xu N, Tsai CL, Lee JT. Transient homologous chromosome pairing marks the onset of X inactivation. Science. 2006;311(5764):1149–52. [PubMed: 16424298]
- 111.
- Xu N, et al. Evidence that homologous X-chromosome pairing requires transcription and Ctcf protein. Nat Genet. 2007;39(11):1390–6. [PubMed: 17952071]
- 112.
- Augui S, et al. Sensing X chromosome pairs before X inactivation via a novel X-pairing region of the Xic. Science. 2007;318(5856):1632–6. [PubMed: 18063799]
- 113.
- Takagi N. Variable X chromosome inactivation patterns in near-tetraploid murine EC x somatic cell hybrid cells differentiated in vitro. Genetica. 1993;88(2-3):107–17. [PubMed: 8224851]
- 114.
- Monkhorst K, et al. The probability to initiate X chromosome inactivation is determined by the X to autosomal ratio and X chromosome specific allelic properties. PLoS One. 2009;4(5):e5616. [PMC free article: PMC2680018] [PubMed: 19440388]
- 115.
- Barr ML, Bertram EG. A morphological distinction between neurones of the male and female and the behaviour of the nucleolar satellite during accelerated nucleoprotein synthesis. Nature. 1949;163(4148):676. [PubMed: 18120749]
- 116.
- Ohno S, Hauschka TS. Allocycly of the X-chromosome in tumors and normal tissues. Cancer Res. 1960;20:541–5. [PubMed: 14428472]
- 117.
- Wutz A, Rasmussen TP, Jaenisch R. Chromosomal silencing and localization are mediated by different domains of Xist RNA. Nat Genet. 2002;30(2):167–74. [PubMed: 11780141]
- 118.
- Chaumeil J, et al. A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes Dev. 2006;20(16):2223–37. [PMC free article: PMC1553206] [PubMed: 16912274]
- 119.
- Clemson CM, et al. The X chromosome is organized into a gene-rich outer rim and an internal core containing silenced nongenic sequences. Proc Natl Acad Sci USA. 2006;103(20):7688–93. [PMC free article: PMC1472506] [PubMed: 16682630]
- 120.
- Goto Y, et al. Differential patterns of histone methylation and acetylation distinguish active and repressed alleles at X-linked genes. Cytogenet Genome Res. 2002;99(1-4):66–74. [PubMed: 12900547]
- 121.
- Heard E, et al. Methylation of histone H3 at Lys-9 is an early mark on the X chromosome during X inactivation. Cell. 2001;107(6):727–38. [PubMed: 11747809]
- 122.
- Mak W, et al. Mitotically stable association of polycomb group proteins eed and enx1 with the inactive x chromosome in trophoblast stem cells. Curr Biol. 2002;12(12):1016–20. [PubMed: 12123576]
- 123.
- Boggs BA, et al. Differentially methylated forms of histone H3 show unique association patterns with inactive human X chromosomes. Nat Genet. 2002;30(1):73–6. [PubMed: 11740495]
- 124.
- Mermoud JE, et al. Histone H3 lysine 9 methylation occurs rapidly at the onset of random X chromosome inactivation. Curr Biol. 2002;12(3):247–51. [PubMed: 11839280]
- 125.
- Peters AH, et al. Histone H3 lysine 9 methylation is an epigenetic imprint of facultative heterochromatin. Nat Genet. 2002;30(1):77–80. [PubMed: 11740497]
- 126.
- Rougeulle C, et al. Differential histone H3 Lys-9 and Lys-27 methylation profiles on the X chromosome. Mol Cell Biol. 2004;24(12):5475–84. [PMC free article: PMC419884] [PubMed: 15169908]
- 127.
- Kohlmaier A, et al. A chromosomal memory triggered by Xist regulates histone methylation in X inactivation. PLoS Biol. 2004;2(7):E171. [PMC free article: PMC449785] [PubMed: 15252442]
- 128.
- Costanzi C, Pehrson JR. Histone macroH2A1 is concentrated in the inactive X chromosome of female mammals. Nature. 1998;393(6685):599–601. [PubMed: 9634239]
- 129.
- Lock LF, Takagi N, Martin GR. Methylation of the Hprt gene on the inactive X occurs after chromosome inactivation. Cell. 1987;48(1):39–46. [PubMed: 3791414]
- 130.
- Norris DP, Brockdorff N, Rastan S. Methylation status of CpG-rich islands on active and inactive mouse X chromosomes. Mamm Genome. 1991;1(2):78–83. [PubMed: 1799791]
- 131.
- Mohandas T, Sparkes RS, Shapiro LJ. Reactivation of an inactive human X chromosome: evidence for X inactivation by DNA methylation. Science. 1981;211(4480):393–6. [PubMed: 6164095]
- 132.
- Russell LB, Montgomery CS. Comparative studies on X-autosome translocations in the mouse. II. Inactivation of autosomal loci, segregation and mapping of autosomal breakpoints in five T (X;1) S. Genetics. 1970;64(2):281–312. [PMC free article: PMC1212403] [PubMed: 5470482]
- 133.
- Duthie SM, et al. Xist RNA exhibits a banded localization on the inactive X chromosome and is excluded from autosomal material in cis. Hum Mol Genet. 1999;8(2):195–204. [PubMed: 9931327]
- 134.
- Keohane AM, et al. H4 acetylation, XIST RNA and replication timing are coincident and define x; autosome boundaries in two abnormal X chromosomes. Hum Mol Genet. 1999;8(2):377–83. [PubMed: 9931347]
- 135.
- Popova BC, et al. Attenuated spread of X-inactivation in an X; autosome translocation. Proc Natl Acad Sci USA. 2006;103(20):7706–11. [PMC free article: PMC1472509] [PubMed: 16679409]
- 136.
- Gartler SM, Riggs AD. Mammalian X-chromosome inactivation. Annu Rev Genet. 1983;17:155–90. [PubMed: 6364959]
- 137.
- Lyon MF. X-chromosome inactivation: a repeat hypothesis. Cytogenet Cell Genet. 1998;80(1-4):133–7. [PubMed: 9678347]
- 138.
- Lyon MF. LINE-1 elements and X chromosome inactivation: a function for "junk" DNA? Proc Natl Acad Sci USA. 2000;97(12):6248–9. [PMC free article: PMC33995] [PubMed: 10841528]
- 139.
- Lyon MF. Do LINEs Have a Role in X-Chromosome Inactivation? J Biomed Biotechnol. 2006;2006(1):59746. [PMC free article: PMC1510946] [PubMed: 16877818]
- 140.
- Lyon MF. No longer all-or-none. Eur J Hum Genet. 2005;13(7):796–7. [PubMed: 15870823]
- 141.
- Furano AV. The biological properties and evolutionary dynamics of mammalian LINE-1 retrotransposons. Prog Nucleic Acid Res Mol Biol. 2000;64:255–94. [PubMed: 10697412]
- 142.
- Waters PD, et al. LINE-1 distribution in Afrotheria and Xenarthra: implications for understanding the evolution of LINE-1 in eutherian genomes. Chromosoma. 2004;113(3):137–44. [PubMed: 15338236]
- 143.
- Parish DA, et al. Distribution of LINEs and other repetitive elements in the karyotype of the bat Carollia: implications for X-chromosome inactivation. Cytogenet Genome Res. 2002;96(1-4):191–7. [PubMed: 12438798]
- 144.
- Wichman HA, et al. Transposable elements and the evolution of genome organization in mammals. Genetica. 1992;86(1-3):287–93. [PubMed: 1334913]
- 145.
- Bailey JA, et al. Molecular evidence for a relationship between LINE-1 elements and X chromosome inactivation: the Lyon repeat hypothesis. Proc Natl Acad Sci USA. 2000;97(12):6634–9. [PMC free article: PMC18684] [PubMed: 10841562]
- 146.
- Carrel L, Willard HF. X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature. 2005;434(7031):400–4. [PubMed: 15772666]
- 147.
- Wang Z, et al. Evidence of influence of genomic DNA sequence on human X chromosome inactivation. PLoS Comput Biol. 2006;2(9):e113. [PMC free article: PMC1557588] [PubMed: 16948528]
- 148.
- Hall LL, et al. Unbalanced X; autosome translocations provide evidence for sequence specificity in the association of XIST RNA with chromatin. Hum Mol Genet. 2002;11(25):3157–65. [PubMed: 12444100]
- 149.
- Sharp AJ, et al. Molecular and cytogenetic analysis of the spreading of X inactivation in X; autosome translocations. Hum Mol Genet. 2002;11(25):3145–56. [PubMed: 12444099]
- 150.
- Sharp A, Robinson DO, Jacobs P. Absence of correlation between late-replication and spreading of X inactivation in an X; autosome translocation. Hum Genet. 2001;109(3):295–302. [PubMed: 11702210]
- 151.
- White WM, et al. The spreading of X inactivation into autosomal material of an x; autosome translocation: evidence for a difference between autosomal and X-chromosomal DNA. Am J Hum Genet. 1998;63(1):20–8. [PMC free article: PMC1377243] [PubMed: 9634520]
- 152.
- Solari AJ, et al. The behavior of sex chromosomes in two human X-autosome translocations: failure of extensive X-inactivation spreading. Biocell. 2001;25(2):155–66. [PubMed: 11590891]
- 153.
- Dobigny G, et al. Viability of X-autosome translocations in mammals: an epigenomic hypothesis from a rodent case-study. Chromosoma. 2004;113(1):34–41. [PubMed: 15243753]
- 154.
- Chureau C, et al. Comparative sequence analysis of the X-inactivation center region in mouse, human and bovine. Genome Res. 2002;12(6):894–908. [PMC free article: PMC1383731] [PubMed: 12045143]
- 155.
- Cantrell MA, Carstens BC, Wichman HA. X chromosome inactivation and Xist evolution in a rodent lacking LINE-1 activity. PLoS One. 2009;4(7):e6252. [PMC free article: PMC2705805] [PubMed: 19603076]
- 156.
- Ke X, Collins A. CpG islands in human X-inactivation. Ann Hum Genet. 2003;67(Pt 3):242–9. [PubMed: 12914576]
- 157.
- Cao R, Zhang Y. SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED-EZH2 complex. Mol Cell. 2004;15(1):57–67. [PubMed: 15225548]
- 158.
- de la Cruz CC, et al. Developmental regulation of Suz 12 localization. Chromosoma. 2005;114(3):183–92. [PubMed: 15986205]
- 159.
- Wang J, et al. Imprinted X inactivation maintained by a mouse Polycomb group gene. Nat Genet. 2001;28(4):371–5. [PubMed: 11479595]
- 160.
- Kalantry S, Magnuson T. The Polycomb group protein EED is dispensable for the initiation of random X-chromosome inactivation. PLoS Genet. 2006;2(5):e66. [PMC free article: PMC1456320] [PubMed: 16680199]
- 161.
- Schoeftner S, et al. Recruitment of PRC1 function at the initiation of X inactivation independent of PRC2 and silencing. Embo J. 2006;25(13):3110–22. [PMC free article: PMC1500994] [PubMed: 16763550]
- 162.
- de Napoles M, et al. Polycomb group proteins Ring1A/B link ubiquitylation of histone H2A to heritable gene silencing and X inactivation. Dev Cell. 2004;7(5):663–76. [PubMed: 15525528]
- 163.
- Fang J, et al. Ring1b-mediated H2A ubiquitination associates with inactive X chromosomes and is involved in initiation of X inactivation. J Biol Chem. 2004;279(51):52812–5. [PubMed: 15509584]
- 164.
- Hernandez-Munoz I, et al. Stable X chromosome inactivation involves the PRC1 Polycomb complex and requires histone MACROH2A1 and the CULLIN3/SPOP ubiquitin E3 ligase. Proc Natl Acad Sci USA. 2005;102(21):7635–40. [PMC free article: PMC1140410] [PubMed: 15897469]
- 165.
- Nusinow DA, et al. Poly(ADP-ribose) polymerase 1 is inhibited by a histone H2A variant, MacroH2A and contributes to silencing of the inactive X chromosome. J Biol Chem. 2007;282(17):12851–9. [PubMed: 17322296]
- 166.
- Kim MY, Zhang T, Kraus WL. Poly(ADP-ribosyl)ation by PARP-1: PAR-laying NAD+ into a nuclear signal. Genes Dev. 2005;19(17):1951–67. [PubMed: 16140981]
- 167.
- Rouleau M, Aubin RA, Poirier GG. Poly(ADP-ribosyl)ated chromatin domains: access granted. J Cell Sci. 2004;117(Pt 6):815–25. [PubMed: 14963022]
- 168.
- Hassa PO, et al. Nuclear ADP-ribosylation reactions in mammalian cells: where are we today and where are we going? Microbiol Mol Biol Rev. 2006;70(3):789–829. [PMC free article: PMC1594587] [PubMed: 16959969]
- 169.
- Kim MY, et al. NAD+-dependent modulation of chromatin structure and transcription by nucleosome binding properties of PARP-1. Cell. 2004;119(6):803–14. [PubMed: 15607977]
- 170.
- Helbig R, Fackelmayer FO. Scaffold attachment factor A (SAF-A) is concentrated in inactive X chromosome territories through its RGG domain. Chromosoma. 2003;112(4):173–82. [PubMed: 14608463]
- 171.
- Ganesan S, et al. BRCA1 supports XIST RNA concentration on the inactive X chromosome. Cell. 2002;111(3):393–405. [PubMed: 12419249]
- 172.
- Silver DP, et al. Further evidence for BRCA1 communication with the inactive X chromosome. Cell. 2007;128(5):991–1002. [PubMed: 17350581]
- 173.
- Vincent-Salomon A, et al. X inactive-specific transcript RNA coating and genetic instability of the X chromosome in BRCA1 breast tumors. Cancer Res. 2007;67(11):5134–40. [PubMed: 17545591]
- 174.
- Xiao C, et al. The XIST noncoding RNA functions independently of BRCA1 in X inactivation. Cell. 2007;128(5):977–89. [PubMed: 17350580]
- 175.
- Blewitt ME, et al. SmcHD1, containing a structural-maintenance-of-chromosomes hinge domain, has a critical role in X inactivation. Nat Genet. 2008;40(5):663–9. [PubMed: 18425126]
- 176.
- Garrick D, et al. Loss of Atrx affects trophoblast development and the pattern of X-inactivation in extraembryonic tissues. PLoS Genet. 2006;2(4):e58. [PMC free article: PMC1440874] [PubMed: 16628246]
- 177.
- Baumann C, De La Fuente R. ATRX marks the inactive X chromosome (Xi) in somatic cells and during imprinted X chromosome inactivation in trophoblast stem cells. Chromosoma. 2009;118(2):209–22. [PMC free article: PMC2877807] [PubMed: 19005673]
- 178.
- Han HJ, et al. SATB1 reprogrammes gene expression to promote breast tumour growth and metastasis. Nature. 2008;452(7184):187–93. [PubMed: 18337816]
- 179.
- Agrelo R, et al. SATB1 defines the developmental context for gene silencing by Xist in lymphoma and embryonic cells. Dev Cell. 2009;16(4):507–16. [PMC free article: PMC3997300] [PubMed: 19386260]
- 180.
- Disteche CM. Escape from X inactivation in human and mouse. Trends Genet. 1995;11(1):17–22. [PubMed: 7900190]
- 181.
- Disteche CM. Escapees on the X chromosome. Proc Natl Acad Sci USA. 1999;96(25):14180–2. [PMC free article: PMC33938] [PubMed: 10588671]
- 182.
- Disteche CM, Filippova GN, Tsuchiya KD. Escape from X inactivation. Cytogenet Genome Res. 2002;99(1-4):36–43. [PubMed: 12900543]
- 183.
- Brown CJ, Greally JM. A stain upon the silence: genes escaping X inactivation. Trends Genet. 2003;19(8):432–8. [PubMed: 12902161]
- 184.
- Looijenga LH, et al. X inactivation in human testicular tumors. XIST expression and androgen receptor methylation status. Am J Pathol. 1997;151(2):581–90. [PMC free article: PMC1858006] [PubMed: 9250171]
- 185.
- Chow JC, et al. Characterization of expression at the human XIST locus in somatic, embryonal carcinoma and transgenic cell lines. Genomics. 2003;82(3):309–22. [PubMed: 12906856]
- 186.
- Migeon BR, et al. Identification of TSIX, encoding an RNA antisense to human XIST, reveals differences from its murine counterpart: implications for X inactivation. Am J Hum Genet. 2001;69(5):951–60. [PMC free article: PMC1274371] [PubMed: 11555794]
- 187.
- Chow JC, et al. Ectopic XIST transcripts in human somatic cells show variable expression and localization. Cytogenet Genome Res. 2002;99(1-4):92–8. [PubMed: 12900550]
- 188.
- Hall LL, et al. An ectopic human XIST gene can induce chromosome inactivation in postdifferentiation human HT-1080 cells. Proc Natl Acad Sci USA. 2002;99(13):8677–82. [PMC free article: PMC124357] [PubMed: 12072569]
- 189.
- Hall LL, et al. X-inactivation reveals epigenetic anomalies in most hESC but identifies sublines that initiate as expected. J Cell Physiol. 2008;216(2):445–52. [PMC free article: PMC3057623] [PubMed: 18340642]
- 190.
- Shen Y, et al. X-inactivation in female human embryonic stem cells is in a nonrandom pattern and prone to epigenetic alterations. Proc Natl Acad Sci USA. 2008;105(12):4709–14. [PMC free article: PMC2290804] [PubMed: 18339804]
- 191.
- Silva SS, et al. X-chromosome inactivation and epigenetic fluidity in human embryonic stem cells. Proc Natl Acad Sci USA. 2008;105(12):4820–5. [PMC free article: PMC2290799] [PubMed: 18339803]
- 192.
- Enver T, et al. Cellular differentiation hierarchies in normal and culture-adapted human embryonic stem cells. Hum Mol Genet. 2005;14(21):3129–40. [PubMed: 16159889]
- 193.
- Hoffman LM, et al. X-inactivation status varies in human embryonic stem cell lines. Stem Cells. 2005;23(10):1468–78. [PubMed: 16123389]
- 194.
- Dhara SK, Benvenisty N. Gene trap as a tool for genome annotation and analysis of X chromosome inactivation in human embryonic stem cells. Nucleic Acids Res. 2004;32(13):3995–4002. [PMC free article: PMC506821] [PubMed: 15284332]
- 195.
- Takagi N. Imprinted X-chromosome inactivation: enlightenment from embryos in vivo. Semin Cell Dev Biol. 2003;14(6):319–29. [PubMed: 15015739]
- 196.
- West JD, et al. Preferential expression of the maternally derived X chromosome in the mouse yolk sac. Cell. 1977;12(4):873–82. [PubMed: 597862]
- 197.
- van den Berg IM, et al. X chromosome inactivation is initiated in human preimplantation embryos. Am J Hum Genet. 2009;84(6):771–9. [PMC free article: PMC2694969] [PubMed: 19481196]
- 198.
- Looijenga LH, et al. Heterogeneous X inactivation in trophoblastic cells of human full-term female placentas. Am J Hum Genet. 1999;64(5):1445–52. [PMC free article: PMC1377883] [PubMed: 10205278]
- 199.
- Ropers HH, Wolff G, Hitzeroth HW. Preferential X inactivation in human placenta membranes: is the paternal X inactive in early embryonic development of female mammals? Hum Genet. 1978;43(3):265–73. [PubMed: 700701]
- 200.
- Daniels R, et al. XIST expression in human oocytes and preimplantation embryos. Am J Hum Genet. 1997;61(1):33–9. [PMC free article: PMC1715845] [PubMed: 9245982]
- 201.
- Ray PF, Winston RM, Handyside AH. XIST expression from the maternal X chromosome in human male preimplantation embryos at the blastocyst stage. Hum Mol Genet. 1997;6(8):1323–7. [PubMed: 9259279]
- 202.
- Bamforth F, Machin G, Innes M. X-chromosome inactivation is mostly random in placental tissues of female monozygotic twins and triplets. Am J Med Genet. 1996;61(3):209–15. [PubMed: 8741865]
- 203.
- Migeon BR, et al. Incomplete X chromosome dosage compensation in chorionic villi of human placenta. Proc Natl Acad Sci USA. 1985;82(10):3390–4. [PMC free article: PMC397781] [PubMed: 3858827]
- 204.
- Migeon BR, Axelman J, Jeppesen P. Differential X reactivation in human placental cells: implications for reversal of X inactivation. Am J Hum Genet. 2005;77(3):355–64. [PMC free article: PMC1226202] [PubMed: 16080112]
- 205.
- Goto T, Wright E, Monk M. Paternal X-chromosome inactivation in human trophoblastic cells. Mol Hum Reprod. 1997;3(1):77–80. [PubMed: 9239711]
- 206.
- Harrison KB. X-chromosome inactivation in the human cytotrophoblast. Cytogenet Cell Genet. 1989;52(1-2):37–41. [PubMed: 2612213]
- 207.
- Harrison KB, Warburton D. Preferential X-chromosome activity in human female placental tissues. Cytogenet Cell Genet. 1986;41(3):163–8. [PubMed: 3956266]
- 208.
- Liu W, Sun X. Skewed X chromosome inactivation in diploid and triploid female human embryonic stem cells. Hum Reprod. 2009;24(8):1834–43. [PubMed: 19429659]
- 209.
- Sato N, et al. Molecular signature of human embryonic stem cells and its comparison with the mouse. Dev Biol. 2003;260(2):404–13. [PubMed: 12921741]
- 210.
- Ginis I, et al. Differences between human and mouse embryonic stem cells. Dev Biol. 2004;269(2):360–80. [PubMed: 15110706]
- 211.
- Xu RH, et al. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nat Methods. 2005;2(3):185–90. [PubMed: 15782187]
- 212.
- Ying QL, et al. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003;115(3):281–92. [PubMed: 14636556]
- 213.
- Boyer LA, et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005;122(6):947–56. [PMC free article: PMC3006442] [PubMed: 16153702]
- 214.
- Brons IG, et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448(7150):191–5. [PubMed: 17597762]
- 215.
- Tesar PJ, et al. New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature. 2007;448(7150):196–9. [PubMed: 17597760]
- 216.
- Rossant J. Stem cells and early lineage development. Cell. 2008;132(4):527–31. [PubMed: 18295568]
- 217.
- Lovell-Badge R. Many ways to pluripotency. Nat Biotechnol. 2007;25(10):1114–6. [PubMed: 17921995]
- 218.
- Xu RH, et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat Biotechnol. 2002;20(12):1261–4. [PubMed: 12426580]
- 219.
- Guo G, et al. Klf4 reverts developmentally programmed restriction of ground state pluripotency. Development. 2009;136(7):1063–9. [PMC free article: PMC2685927] [PubMed: 19224983]
- 220.
- Bao S, et al. Epigenetic reversion of post-implantation epiblast to pluripotent embryonic stem cells. Nature. 2009;461(7268):1292–5. [PMC free article: PMC3863718] [PubMed: 19816418]
- 221.
- Hanna J, et al. Metastable pluripotent states in NOD-mouse-derived ESCs. Cell Stem Cell. 2009;4(6):513–24. [PMC free article: PMC2714944] [PubMed: 19427283]
- 222.
- Nichols J, Smith A. Naive and primed pluripotent states. Cell Stem Cell. 2009;4(6):487–92. [PubMed: 19497275]
- 223.
- Zvetkova I, et al. Global hypomethylation of the genome in XX embryonic stem cells. Nat Genet. 2005;37(11):1274–9. [PubMed: 16244654]
- 224.
- Lagarkova MA, et al. Diverse epigenetic profile of novel human embryonic stem cell lines. Cell Cycle. 2006;5(4):416–20. [PubMed: 16479162]
- 225.
- Rugg-Gunn PJ, Ferguson-Smith AC, Pedersen RA. Epigenetic status of human embryonic stem cells. Nat Genet. 2005;37(6):585–7. [PubMed: 15864307]
- 226.
- Allegrucci C, et al. Restriction landmark genome scanning identifies culture-induced DNA methylation instability in the human embryonic stem cell epigenome. Hum Mol Genet. 2007;16(10):1253–68. [PubMed: 17409196]
- X Chromosome Inactivation and Embryonic Stem Cells - Madame Curie Bioscience Dat...X Chromosome Inactivation and Embryonic Stem Cells - Madame Curie Bioscience Database
- NOTCH SIGNALING AND THE DEVELOPING INNER EAR - Madame Curie Bioscience DatabaseNOTCH SIGNALING AND THE DEVELOPING INNER EAR - Madame Curie Bioscience Database
- Restriction-Modification Systems as Mobile Epigenetic Elements - Madame Curie Bi...Restriction-Modification Systems as Mobile Epigenetic Elements - Madame Curie Bioscience Database
- Molecular Mechanisms of ErbB2-Mediated Breast Cancer Chemoresistance - Madame Cu...Molecular Mechanisms of ErbB2-Mediated Breast Cancer Chemoresistance - Madame Curie Bioscience Database
- The Role of Neuropilin in Vascular and Tumor Biology - Madame Curie Bioscience D...The Role of Neuropilin in Vascular and Tumor Biology - Madame Curie Bioscience Database
Your browsing activity is empty.
Activity recording is turned off.
See more...