

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Coffin JM, Hughes SH, Varmus HE, editors. Retroviruses. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press; 1997.
Retroviral particles usually are purified on the basis of their size and density. Because the particles are shed into the extracellular medium, purification is simple and efficient if disruption of cells can be avoided, as, for example, when the virus is derived from cultured mammalian or avian cells or from the plasma of an animal. The virus first is collected by centrifugation of the growth medium. The resulting pellet is redissolved and the particles are sedimented to equilibrium, usually by centrifugation in a gradient of sucrose. The density of retroviruses is approximately 1.16 g/ml, corresponding to about 35% w/w sucrose. In favorable cases, for example, Rous sarcoma virus (RSV)-infected chick embryo fibroblasts, as much as 1% of the total cell protein synthesis is devoted to viral proteins, and such cells can release up to several micrograms of virus per milliliter of medium. However, typical yields may be considerably lower. Virus purified in this way invariably is contaminated with some cellular proteins, because membrane vesicles from broken or intact cells have a density similar to that of the virus. Higher levels of purification can be achieved by selecting not only for particle density, but also for particle size, for example, by rate zonal sedimentation. The most concentrated and clean source of any retrovirus is the plasma of chicks infected with avian myeloblastosis virus (AMV), which can contain as much as several milligrams of virus per milliliter of plasma (1 mg is about 1012 virions). It is for this reason that many of the early biochemical studies of viral structural proteins and of reverse transcriptase were carried out with this member of the ASLV genus. Like almost all stocks of highly oncogenic retroviruses, AMV is in fact a mixture of the virus carrying the oncogene (here, myb) and associated helper viruses, in this case called myeloblastosis-associated viruses (MAVs). MAVs are structurally indistinguishable from avian leukemia viruses (ALVs).
As with many other types of viruses, not all of the virions in a preparation are infectious. Typically for retroviruses, the ratio of physical to infectious particles is 100:1 or greater. Freshly harvested virus is the most infectious, with the titer of newly shed virus in the medium decreasing with a half-life in the range of a few hours at 37oC. Thus, in most biochemical or cell biological studies, the measurements in fact reflect the properties of the inactive particles, since these are the predominant population. Interpretations that do not take cognizance of the vast excess of inactive virions may be flawed. Infectious particles are rapidly inactivated by standard disinfecting treatments, for example, detergents. These facts have obviously important implications for those retroviruses that cause animal or human diseases.
According to early analyses, retroviral particles are composed of about 1–2% RNA, about 35% lipids, and about 65% protein. By mass, perhaps two thirds of the RNA in particles is the viral genomic RNA, with the remainder being accounted for largely by transfer RNAs, as well as other small host RNAs. Although the viral envelope is derived from the cell plasma membrane by budding, its composition differs somewhat from that of the plasma membrane, being enriched in sphingomyelin and cholesterol (Quigley et al. 1971; Aloia et al. 1993). In clean preparations of virus, about three quarters of the total protein component is due to the internal structural proteins, the Gag proteins. Most of the remainder is composed of viral envelope glycoproteins (Env protein) and to a lesser extent the viral enzymes reverse transcriptase, integrase, and protease, and some cellular proteins. Even the purest viral preparation shows a panoply of unidentified minor polypeptides by SDS-polyacrylamide gel electrophoresis if sensitive methods of detection are used. Many if not most of these may become incorporated adventitiously during assembly, but, as discussed below, some proteins with known or suspected function in the viral life cycle have been identified.
Particle size for viruses typically is measured either by electron microscopy or by rate zonal sedimentation. As usually applied, neither method is very accurate. Thin-section techniques require harsh fixation, and the appearance of the particles finally visualized depends on the plane of sectioning. Negative staining can cause deformations in particles. Sedimentation, for example, in sucrose gradients, is rather insensitive to size, with a doubling in size resulting in only about 30% change in sedimentation rate. In addition, several assumptions must be made to allow calculation of the size of a particle from its sedimentation rate. In thin-section electron microscopy, retroviruses measure about 80–120 nm in diameter. Since very few studies have compared different viruses in the same experiment, it is uncertain if size differs among the retroviral genera or if the spread in measured diameters represents other factors. In rate zonal sedimentation, viral particles sediment at about 600S.
Morphology, as defined by electron microscopy, is one of the major criteria for classification of viruses. The technique of negative staining shows the perimeter and sometimes the center of the virion outlined by the surrounding accumulation of heavy metal, typically uranyl acetate or phosphotungstate (Fig. 1I). In this rapid procedure, virus is simply adsorbed to a coated grid and then exposed briefly to the solution of heavy metal before viewing. For structures that can be penetrated by the heavy metal, negative staining usually allows excellent visualization of structural details, but the lipid membrane of enveloped viruses typically is not penetrated by the stain, limiting the usefulness of this technique for retroviruses. For many purposes, the thin-sectioning technique provides more information, but it requires fixation of virus or cells by a crosslinking reagent, dehydration, staining, and embedding into plastic before sectioning and viewing.
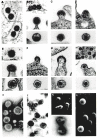
By the early 1970s, several distinctive morphologies of retroviral particles had been identified by thin-section techniques (Bernhard 1958; deHarven 1974). Type-A particles were initially defined as the nonenveloped cytoplasmic precursor particles to type-B (Fig. 1B) and type-D retroviruses (Fig. 1E). Alternatively, they have been defined by the prototypic “intracisternal type-A particles” (IAPs), which bud into and accumulate in the endoplasmic reticulum of a number of normal and transformed rodent cells (Fig. 1A). IAPs are expressed from endogenous retrovirus-like elements that are present in multiple copies in rodent genomes (Chapter 8). Unlike the precursor particles in the cytoplasm, IAPs carry an envelope because they have budded through a membrane (although most lack envelope proteins). The characteristic feature of type-A particles is an electronlucent center and one or two concentric electrondense rings. Thus, in thin sections, these particles have the appearance of a doughnut. It is a useful and now common practice to call this morphology “immature,” because mutant retroviruses unable to process their internal proteins proteolytically give rise to particles with similar morphologies (see below and Chapter 7). Indeed, the Gag protein is unprocessed in IAPs as well as in cytoplasmic precursor particles. Since proteolysis follows assembly, even wild-type infectious viral particles in the process of budding show the same characteristic structure, before they have been released and become mature by the condensation of the core.
Mature, infectious retroviral particles have a condensed core of electrondense material. Many of them are of type-C morphology (Fig. 1C,D), with a round or sometimes slightly angular core centered in the middle of the particle. Examples include the MLVs and ASLVs. Type-B viruses show an eccentrically placed round core, with mouse mammary tumor virus (MMTV) being the prototype (Fig. 1B). Type-D viruses like Mason-Pfizer monkey virus (M-PMV) have a bar-shaped core (Fig. 1E). Type-B and type-D viruses as well as spumaviruses are distinguished by their mode of assembly. Whereas most retroviruses assemble their cores at the plasma membrane prior to envelopment and release (Fig. 1C,F,G), these viruses assemble their cores in the cytoplasm. The fully assembled core structures are then transported to the plasma membrane for envelopment, release, and maturation (Chapter 7). Some mature retroviruses have a distinctive morphology that has not been given a letter name. For example, although in budding, the lentiviruses (prototype HIV-1) resemble type-C viruses, the core in the mature virion has the shape of a cone (Fig. 1G). Depending on the orientation of the plane of sectioning, the core may thus take on different appearances.
The structural features that underlie the different morphologies of retroviral particles and the apparently different assembly routes are not well understood. A number of cases have been reported for which virus-like particles can assemble from chimeric viral Gag proteins, for example, part HIV-1 and part ALV. These techniques may make it possible to pinpoint the structural determinants responsible for the characteristic morphology of each virus type with a graded series of chimeras. These determinants could be quite small. In the case of M-PMV, a single point mutation in the Gag protein is able to change the route of assembly so that the mutant now resembles a type-C retrovirus (Rhee and Hunter 1990). Which of the several proteolytic cleavages in Gag are responsible for the “collapse” of the core is not yet certain. Systematic mutations at each of the cleavage sites in the Gag protein, coupled with thin-section electron microscopy, may make it possible to decide which cleavages are central to the morphological changes seen upon maturation. It is not excluded that morphological maturation is a separate step, not invariably following proteolytic cleavages of Gag. This notion is suggested by the observation that certain gross mutations in the nucleocapsid domain of Gag prevent maturation, despite nearly normal levels of proteolytic processing (Berkowitz et al. 1995b).
Both negative staining and thin-section techniques show projections emanating from the viral envelope, which comprise the viral envelope glycoproteins (Chapter 3). The number of such projections varies greatly among different viruses, and even among different strains or isolates of the same virus. This variability is poorly understood and often is attributed to the propensity of the surface glycoprotein to fall off spontaneously during purification or storage. However, in some cases, even freshly isolated viruses show few projections, and these viral preparations also contain little envelope protein by biochemical measurements. In contrast, some retroviruses, for example, spumaviruses, typically are densely studded with glycoproteins (Fig. 1H). In ASLV, each projection seen in the electron microscope is probably a trimer, and in HIV, conflicting measurements have suggested either a dimer or a tetramer.
More refined electron microscopic techniques are being applied to retroviruses and in the future may yield a more detailed understanding of structure. By scanning electron microscopy, the viral particle appears to be three dimensional, with a rough surface presumably corresponding to the envelope spikes (Fig. 1L). The budding process can be readily visualized by this technique. In thin sections, the three-dimensional appearance of objects can sometimes be reconstructed if the section is viewed at different angles. With information processed from tilted sections of HIV-1 cores, it has been possible to create a model of the form of the core as it is preserved after fixation and staining (Höglund et al. 1992). Electron microscopic thin-section techniques suffer from the inherent problems of fixation and staining. Since harsh conditions are used to prepare the sections, the structures seen are only those that have been preserved by crosslinking. Cryoelectron microscopy (cryo-EM) obviates these problems, since the virus is observed directly as an unstained particle in a thin sheet of noncrystalline ice at the temperature of liquid nitrogen. Although such images have low contrast, if the virus has detectable symmetry (e.g., icosahedral symmetry), computer-assisted averaging can be used to construct a three-dimensional image of the virus. Detailed high-resolution reconstructions have been published for numerous spherical viruses, including the alphaviruses (Cheng et al. 1995; Fuller et al. 1995), which, as simple enveloped RNA viruses, can serve as models for retroviruses. Cryo-EM analyses of retroviruses have been reported only recently for mature and immature MLVs (E. Kubalek et al., in prep.) and for HIV-like Gag particles expressed in insect cells (S.D. Fuller et al., in prep.). Immature particles show a spoke-like structure inside the envelope for both HIV and MLV (Fig. 2), as also occasionally seen in negative-stain electron micrographs. Mature particles of MLV simply show a spherical core, with no obvious symmetrical features (Fig. 2A).
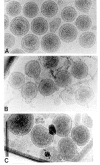
A feature of retroviral structure common to many published models is the depiction of the mature or the immature core as icosahedral. Although there are theoretical considerations as well as abundant precedents from other viruses that make such a shape likely a priori, direct evidence remains sketchy. The mature core for the lentiviruses and type-D viruses is not even approximately spherical. For type C and probably most retroviruses, mature cores can be isolated only in very poor yield, because agents that remove the lipid envelope also cause the core to dissociate, and thus negative-stain techniques are difficult to apply. (Equine infectious anemia virus [EIAV] seems to be an exception, since stable cores can be readily isolated by simple detergent treatment of virions (Roberts and Oroszlan 1989).) Nevertheless, negative staining and freeze-etching do occasionally suggest icosahedral symmetry for C-type viruses (Nermut et al. 1972). Icosahedral symmetry of Env proteins on the surface of rare HIV virions has also been observed (Gelderblom 1991). On the other hand, some investigators have interpreted thin sections of retroviruses to suggest a helical core (Sarkar et al. 1971). Analyses of MLV by cryo-EM (E. Kubalek et al., in prep.) do not provide evidence for an icosahedral shape of the core.
In contrast to mature cores, immature cores are quite stable to weak detergents, and thus can be isolated readily and studied by negative-staining techniques, for example, from mutant viruses with a defective protease (Chapter 6). The relative stability of the immature core compared with the fragility of the mature core with its processed proteins may reflect the different structural demands of assembly and disassembly. However, even in immature cores, native structure may be compromised by detergent (S.D. Fuller et al., in prep.). Negative-stain electron microscopy of immature, delipidated HIV particles shows evidence of a hexagonal arrangement of subunits in local areas (Nermut et al. 1994), but this does not necessarily imply icosahedral symmetry. Other microscopic evidence for an icosahedral core in HIV has also been presented but is only suggestive (Nermut et al. 1993). The fact that cryo-EM pictures of immature particles do not reveal clear icosahedral symmetry in either MLV (E. Kubalek et al., in prep.) or HIV (S.D. Fuller et al., in prep.) suggests that if such symmetry features exist, they may be unstable after virion formation. An alternative model for the structure of immature retroviral particles postulates a helical symmetry (Campbell and Vogt 1995), as envisioned in early studies on MLV (Yoshinaka and Luftig 1977). A more complete understanding of the molecular structure of retroviral particles thus may have to await the development of new techniques, for example, computer programs to help interpret the cryo-EM images of nonicosahedral viruses. A different approach that has been successfully used in other viral systems is reconstitution of virus-like particles from highly purified proteins obtained by expression in Escherichia coli. The first studies on in vitro assembly of particles from purified, expressed retroviral proteins have been reported recently (Campbell and Vogt 1995 and unpubl.; Klikova et al. 1995; Sakalian et al. 1996).
The exact juxtaposition of the components of the retroviral particle remains uncertain, and hence models are necessary to represent the structure of the virion. The most useful models represent in a pictorial form what is known about the topographic relationships of components of the particle, as well as other aspects of their structure and function. Predictions may also be incorporated into models if appropriate. Numerous models for virions have been published, some highly detailed, but all are based to some extent on conjecture and analogy with other viruses. One of the earliest and perhaps most influential, presented in 1978 (Bolognesi et al. 1978), presaged much of what was learned later from biochemical studies about the internal organization of the virion. Modern versions of this model that incorporate newer information about structural proteins are shown in Figure 3. The drawing is for HIV-1, but with minor modifications, it can apply to other retroviruses as well. Some of the details of the model are discussed at the end of this chapter.
- Purification, Composition, and Morphology of Virions - RetrovirusesPurification, Composition, and Morphology of Virions - Retroviruses
Your browsing activity is empty.
Activity recording is turned off.
See more...