Macrolides-Lincosamides-Streptogramins-Ketolides
Macrolides, lincosamides, and streptogramins (MLS) are recommended as alternate antibiotics for the treatment of S. pyogenes infections in patients who are allergic to β-lactams or in cases of penicillin failure (Bisno, Gerber, Gwaltney, Jr., Kaplan, & Schwartz, 2002). Although MLS compounds are chemically distinct, they are considered to be a unique group, since they share a similar mode of action through binding to (or in the vicinity of) the ribosomal peptidyl transferase center (PTC) (Schlünzen, et al., 2001). Also, they present an overlapping spectrum of activity within the same MIC ranges []. Practically, macrolides are classified according to the number of atoms that form the lactone ring, and there are 14- (e.g. erythromycin or clarithromycin), 15- (e.g. azithromycin), or 16-membered macrolides (e.g. spiramycin or josamycin). Notably, a new class of molecules has been recently developed that have evolved from macrolides, and these are known as ketolides (e.g. telithromycin). Lincosamides comprise only two members: lincomycin and its semisynthetic derivative, clindamycin. Streptogramins are actually composed of a mixture of two compounds that act synergistically: streptogramin A (e.g. dalfopristin) and streptogramin B (e.g. quinupristin) (Dang, Nanda, Cooper, Greenfield, & Bronze, 2007).
The first macrolide-resistant S. pyogenes isolate was reported in the USA in 1968 (Sanders, Foster, & Scott, 1968). In the late 1990s and early 2000s, the incidence of macrolide resistance dramatically increased in several European countries, such as Finland, France, Greece, Italy, Portugal, and Spain, with erythromycin resistance rates usually exceeding 20% (Seppälä, et al., 1997; Granizo, Aguilar, Casal, Dal-Ré, & Baguero, 2000; Bingen, et al., 2004; Silva-Costa, Ramirez, & Melo-Cristino, 2005; Creti, et al., 2007; Richter, et al., 2008). Despite some geographical variations, rates of macrolide resistance in the USA have remained relatively low (around 5%) since the 1990s (Brown & Rybak, 2004; Tanz, et al., 2004; Richter, et al., 2005). More recently, some European studies have described a significant decrease in erythromycin resistance in S. pyogenes, such as in France or Spain (d'Humières, et al., 2012; Montes M. , et al., 2014). From an epidemiological point of view, it was frequently demonstrated that the prevalence of erythromycin-resistant S. pyogenes correlated well with the total consumption of macrolide antibiotics in particular geographical areas (Seppälä, et al., 1997; Granizo, Aguilar, Casal, Dal-Ré, & Baguero, 2000; Seppälä, Klaukka, Lehtonen, Nenonen, & Huovinen, 1995; Albrich, Monnet, & Harbarth, 2004; Bergman, et al., 2004; Hsueh, Shyr, & Wu, 2005). Besides antibiotic consumption, changes in the clonal composition of the S. pyogenes population may also be an important cause for fluctuations in macrolide resistance rates (Montes, Tamayo, Mojica, García-Arenzana, Esnal, & Pérez-Trallero, 2014; Silva-Costa, Friães, Ramirez, & Melo-Cristino, 2012).
Notably, a relationship between virulence and macrolide resistance has emerged. Specifically, erythromycin resistance is associated with the increased cell invasiveness of S. pyogenes (Facinelli, Spinaci, Magi, Giovanetti, & Varaldo, 2001). Indeed, this association could be due to the presence of the prtF1 gene, which is more frequently found among macrolide-resistant strains. The prtF1 gene encodes the fibronectin-binding protein F1, an adhesion that allows S. pyogenes to be efficiently internalized by and survive within human respiratory cells (Facinelli, Spinaci, Magi, Giovanetti, & Varaldo, 2001; Haller, Fluegge, Arri, Adams, & Berner, 2005). By contrast, biofilm formation could be an important factor to explain therapeutic failures and recurrences due to macrolide-susceptible S. pyogenes clinical isolates (Baldassarri, et al., 2006). Additionally, erythromycin-susceptible strains form a significantly thicker biofilm than resistant isolates, while those harboring erm-class genes have a less organized biofilm than mef(A)-positive strains (see below). Finally, the presence of prtF1 appears to be negatively associated with the ability to form biofilm (Baldassarri, et al., 2006).
MLS resistance may be due to several different mechanisms: (i) post-transcriptional target site modifications caused by rRNA methylases (erm-class genes); (ii) target mutations (in 23S rRNA or ribosomal proteins L4 and L22); or (iii) the acquisition of active efflux (mef genes) (Leclercq & Courvalin, 1991; Weisblum, 1995a).
All genes of the erm family encode methyltransferases that specifically add one or two methyl residues on the N6 amino group of the highly conserved adenine A2058 in domain V of the 23S rRNA, which corresponds to the ribosomal PTC (Lai & Weisblum, 1971). This ribosomal methylation conveys cross-resistance to macrolides-lincosamides-streptogramins B (MLSB) that can be constitutively expressed (the so-called cMLSB phenotype) or inducibly expressed (the so-called iMLSB phenotype) [] (Weisblum, 1995b). In streptococci, resistance is commonly mediated by two classes of methylases encoded by erm(B) and erm(TR) genes, and both are usually located chromosomally (Leclercq, 2002). The erm(B) gene (previously known as ermAM) was first identified in Streptococcus sanguinis (Horinouchi, Byeon, & Weisblum, 1983). Because erm(TR) shows 82.5% nucleotide identity with the original erm(A) gene (Seppälä, Skurnik, Soini, Roberts, & Huovinen, 1998), it has been proposed that erm(TR) belongs to the erm(A) class (Roberts, et al., 1999). However, the erm(A) subclass erm(TR) will be referred to as erm(TR) throughout this work.
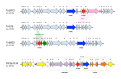
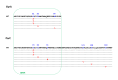
Common phenotypes of MLS resistance in S. pyogenes. (A) Wild-type susceptible strain (S. pyogenes ATCC 19615); (B) S. pyogenes containing an erm(B) gene constitutively expressed; (C) S. pyogenes containing an erm(TR) gene inducibly expressed (a D-shaped (more...)
Whereas erm(B) is primarily associated with a cMLSB phenotype and rarely with a iMLSB phenotype, macrolide resistance conferred by erm(TR) in streptococci is usually expressed inducibly even if some cMLSB
erm(TR)-positive strains have occasionally been reported [] (Leclercq, 2002). In S. pyogenes, there is a substantial heterogeneity of susceptibility patterns among inducibly-resistant isolates, which have been subdivided into three distinct subtypes, designated i-MLSB-A, iMLSB-B, and iMLSB-C [] (Arpin, Canron, Noury, & Quentin, 1999; Giovanetti, Montanari, Mingoia, & Varaldo, 1999; Giovanetti, Montanari, Marchetti, & Varaldo, 2000; Betriu, et al., 2000; Malbruny, et al., 2002; Bingen, et al., 2002). In contrast to cMLB isolates that are highly resistant to lincosamides, all types of iMLSB are associated with susceptibility to lincosamides [ and ]. Phenotypically, iMLSB-A strains are highly resistant to all macrolides; iMLSB-B strains are highly resistant to 14- and 15-membered macrolides, but are susceptible to 16-membered macrolides; and iMLSB-C strains present low-level resistance to 14- and 15-membered macrolides, but remain susceptible to 16-membered macrolides []. cMLSB and iMLSB-A isolates usually harbor the erm(B) gene, while iMLSB-B and iMLSB-C isolates possess the erm(TR) gene (Giovanetti, Montanari, Mingoia, & Varaldo, 1999). Notably, resistance to ketolides is observed in cMLSB and iMLSB-A S. pyogenes isolates, while resistance is correlated to the degree of dimethylation by Erm(B) of the A2058 of the 23S rRNA (Douthwaite, Jalava, & Jakobsen, 2005). Even though the Erm(TR) methylase in S. pyogenes is not inducible with lincosamides, clindamycin resistance (MIC, 64 mg/L) due to constitutive expression can easily be obtained in vitro (at a frequency of ca. 10-7) from inducible erm(TR)-harboring strains [] (Fines, Gueudin, Ramon, & Leclercq, 2001). This is due to alterations in the structure of regulatory sequences, which are composed of two leader peptides (15 and 19 amino acids) involved in post-transcriptional regulation (the so-called translational attenuation) that controls the expression of the methylase gene, in a manner similar to that found for erm(C) in staphylococci (Weisblum, 1995b; Horinouchi & Weisblum, 1980). These modifications in the promoter sequence (including base changes, insertions/duplications, or deletions) have also been described in clinical isolates (Doktor & Shortridge, 2005; Malhotra-Kumar, et al., 2009). Mechanistically, these modifications are responsible for modifications in the mRNA secondary structure that result in changes in accessibility of the ribosome-binding site and the initiation codon of the methylase to the ribosomes, and subsequently for the translation of the erm(TR) transcripts [] (Fines, Gueudin, Ramon, & Leclercq, 2001; Doktor & Shortridge, 2005; Malhotra-Kumar, et al., 2009).
Profiles of susceptibility to MLS antibiotics in S. pyogenes according to phenotypes/ genotypes of resistance (Arpin, Canron, Noury, & Quentin, 1999; Giovanetti, Montanari, Mingoia, & Varaldo, 1999; Giovanetti, Montanari, Marchetti, & (more...)
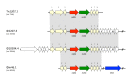
Unusual phenotypes of MLS resistance in S. pyogenes. (A) Wild-type susceptible strain (S. pyogenes ATCC 19615); (B) S. pyogenes containing an erm(TR) gene constitutively expressed due to attenuator alterations (Fines, Gueudin, Ramon, & Leclercq, (more...)
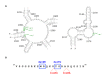
Predicted mRNA secondary structures of the erm(TR) regulatory region (called the attenuator). (A) Conformational isomer of the wild-type erm(TR) mRNA (Malhotra-Kumar, et al., 2009). Open reading frames (ORFs) are indicated in blue: LP1, leader peptide (more...)
Originally identified as part of Tn917 on a non-conjugative plasmid from E. faecalis (Tomich, An, & Clewell, 1980), the erm(B) gene in S. pyogenes is carried by different elements, depending on whether it is expressed constitutively or inducibly (Varaldo, Montanari, & Giovanetti, 2009). When constitutively expressed, this gene is carried by Tn916 family elements, such as Tn3872 (ca. 24 kb) or Tn6002 (ca. 21 kb) [] (Varaldo, Montanari, & Giovanetti, 2009; Brenciani, Bacciaglia, Vecchi, Vitali, Varaldo, & Giovanetti, 2007; Brenciani, Tiberi, Morici, Oryasin, Giovanetti, & Varaldo, 2012; Brenciani, Tiberi, Morroni, Mingoia, Varaldo, & Giovanetti, 2014). Tn3872 results from the insertion of Tn917 into orf9 of Tn916, with erm(B) thus physically linked to tet(M), while conjugal transfer has been demonstrated from S. pyogenes to S. pyogenes [] (Brenciani, Bacciaglia, Vecchi, Vitali, Varaldo, & Giovanetti, 2007). Tn6002 corresponds to the insertion of the erm(B) element (ca. 3 kb) between orf20 and orf19 of Tn916, which also leads to an erm(B)/tet(M) linkage, and for which intraspecific conjugal transfer has been demonstrated in S. pyogenes (Brenciani, Bacciaglia, Vecchi, Vitali, Varaldo, & Giovanetti, 2007). When inducibly expressed, erm(B) is carried by an element originally named Tn1116 (ca. 48 kb), but now renamed ICESp1116, since it has been demonstrated to belong to the TnGBS family of integrative and conjugative elements (ICEs) (Brenciani, Bacciaglia, Vecchi, Vitali, Varaldo, & Giovanetti, 2007; Brenciani, Tiberi, Morici, Oryasin, Giovanetti, & Varaldo, 2012; Brenciani, Tiberi, Morroni, Mingoia, Varaldo, & Giovanetti, 2014). This element presents a unique mosaic structure related to the TnGallo1 from Streptococcus gallolyticus with two inserted fragments separated by an IS1216: the erm(B)-containing fragment (derived from the plasmid pSM19035) and the right-hand portion of Clostridium difficile Tn5397 that contains a truncated tet(M) gene [] (Brenciani, Tiberi, Morici, Oryasin, Giovanetti, & Varaldo, 2012).
erm(B)-carrying genetic elements (Varaldo, Montanari, & Giovanetti, 2009; Brenciani A. , et al., 2007; Brenciani A. , et al., 2012; Brenciani A. , et al., 2014). Open reading frames (ORFs) are indicated by horizontal arrows; erm(B) and tet(M) (more...)
Different erm(TR)-carrying ICE elements have been described in S. pyogenes: ICE 10750-RD.2 (ca. 49 kb) and ICESp1108 (ca. 45 kb) in tetracycline-susceptible strains, and ICESp2905 (ca. 66 kb) in tetracycline-resistant strains, due to the presence of the tet(O) gene [] (Varaldo, Montanari, & Giovanetti, 2009; Brenciani, Tiberi, Bacciaglia, Petrelli, Varaldo, & Giovanetti, 2011; Giovanetti, Brenciani, Tiberi, Bacciaglia, & Varaldo, 2012). While the element ICE 10750-RD.2 is integrated into an hsdM chromosomal gene-encoding host DNA restriction/modification methyltransferase, both ICESp1108 and ICESp2905 are integrated in the chromosome at the 3’ end of the conserved RNA uracil methyltransferase (rum) gene (Varaldo, Montanari, & Giovanetti, 2009; Brenciani, Tiberi, Bacciaglia, Petrelli, Varaldo, & Giovanetti, 2011). The ICESp2905 results from one ICE (ICESp2907) being integrated into another (ICESp2906), with the former containing erm(TR) and the latter containing tet(O), and the whole inserted into a scaffold of clostridial origin [] (Giovanetti, Brenciani, Tiberi, Bacciaglia, & Varaldo, 2012). Notably, all these structures share an almost identical conserved core sequence (ca. 2 kb) that includes erm(TR) and two adjacent antibiotic resistance (tetronasin and spectinomycin) genes [] (Brenciani, Tiberi, Bacciaglia, Petrelli, Varaldo, & Giovanetti, 2011). Note that erm(TR) can be transferred by conjugation to susceptible recipients of S. pyogenes and other Gram-positive bacteria (Giovanetti, et al., 2002). In addition, it has been shown that Peptostreptococcus spp. may serve as an important reservoir for erm(TR)-mediated macrolide resistance (Reig, Galan, Baquero, & Perez-Diaz, 2001).
erm(TR)-carrying genetic elements that depend on tetracycline susceptibility/resistance (Varaldo, Montanari, & Giovanetti, 2009; Brenciani A. , et al., 2011; Giovanetti E. , Brenciani, Tiberi, Bacciaglia, & Varaldo, 2012). Open reading (more...)
Another methylase gene, erm(T) (previously named ermGT), has also been identified in S. pyogenes that expresses an iMLSB resistance phenotype (Woodbury, et al., 2008). Originally described in Lactobacillus reuteri in a chromosomal location (Tannock, et al., 1994), this gene is borne on a small mobilizable plasmid (ca. 5 kb) in S. pyogenes (Woodbury, et al., 2008; DiPersio, DiPersio, Beach, Loudon, & Fuchs, 2011).
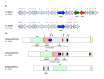
Macrolide resistance can also be due to ribosomal mutations, either in the domain V of 23S rRNA (rrn) operons or in the ribosomal proteins L4 and L22 (rplD and rplV genes, respectively) (Leclercq, 2002). In S. pyogenes, two clinical isolates with such target mutations were described for the first time in 2002 (Malbruny, et al., 2002). The former strain exhibited a C2611U mutation in the domain V of all six 23S rRNA copies, and was phenotypically resistant to azithromycin and clindamycin, but remained susceptible to erythromycin and spiramycin [ and , ] (Malbruny, et al., 2002). The latter strain harbored an insertion of six nucleotides in the rplD gene sequence, which resulted in a KG insertion after position 69 in the L4 ribosomal protein, and was phenotypically resistant to azithromycin and spiramycin, but remained susceptible to erythromycin (borderline) and clindamycin [ and , ] (Malbruny, et al., 2002). Since then, additional ribosomal mutations in the L4 ribosomal protein have been reported in a few clinical isolates []: a two-amino-acid deletion (65RW66), an RA insertion after position 73, and a TG deletion at positions 70 to 71 (Bingen, et al., 2002; Bozdogan, Appelbaum, Ednie, Grivea, & Syrogiannopoulos, 2003). Other mutations in 23S rRNA have also been identified in clinical isolates []: a A2058G mutation (5/6 mutated rrn copies) with resistance to all macrolides, ketolides, and lincosamides (Jalava, Vaara, & Huovinen, 2004); A2058G (n=4) and A2059G (n=2) mutations in clinical isolates that present a cMLSB phenotype (Tanz, et al., 2004; Richter, et al., 2005); dual mutations (A2058G and U2166C) in seven clonally-related strains resistant to macrolides and ketolides (Farrell, Shackcloth, Barbadora, & Green, 2006); and in two isolates with a A2058G substitution with a cMLSB phenotype (Montes, Tamayo, Mojica, García-Arenzana, Esnal, & Pérez-Trallero, 2014).
Ribosomal mutations responsible for MLS resistance in S. pyogenes. (A) Secondary structure of the domain V of 23S rRNA. Positions (according to E. coli numbering) and mutations described in S. pyogenes are indicated in green (Tanz, et al., 2004; Richter, (more...)
Active efflux due to mef genes is responsible for resistance only to 14- and 15-membered macrolides (the so-called M phenotype), while there is no resistance to 16-membered macrolides, lincosamides, and streptogramins (Sutcliffe, Tait-Kamradt, & Wondrack, 1996). Mef proteins are proton-dependent efflux pumps that belong to the major facilitator superfamily (MFS) with 12 transmembrane segments (TMS) (Poole, 2005). Several allelic variants or subclasses of the mef gene have been described, mainly mef(A) originally reported in S. pyogenes (Clancy, et al., 1996) and mef(E) later identified in S. pneumoniae (Tait-Kamradt, et al., 1997). Because these two mef genes show 90% nucleotide identity, it has been proposed to consider them as a single class, designated mef(A) (Roberts, et al., 1999). However, some authors argued for the need to distinguish these determinants, since there are major differences between them (Klaassen & Mouton, 2005). First, erythromycin MICs associated with the mef(A) gene (MIC50, 16 μg/ml) are higher than those associated with mef(E) (MIC50, 8 μg/ml) (Amezaga, Carter, Cash, & McKenzie, 2002; Blackman Northwood, et al., 2009). Second, and most importantly, these determinants are carried by completely different genetic elements (Del Grosso, et al., 2002). Note that the mef(A) subtype is by far the most prevalent mef allele among S. pyogenes clinical isolates (Blackman Northwood, et al., 2009; Sangvik, Littauer, Simonsen, Sundsfjord, & Dahl, 2005; Ardanuy, et al., 2005).
In S. pneumoniae, the mef(A) gene is part of Tn1207.1 (ca. 7 kb), a 7.2-kb defective transposon integrated into the chromosome, while the mef(E) is borne by a transferable macrolide efflux genetic assembly (mega) element (ca. 5 kb) that is integrated into composite transposons Tn2009 (ca. 23 kb) or Tn2010 (ca. 26 kb) (Varaldo, Montanari, & Giovanetti, 2009). In S. pyogenes, the mef(A) gene is carried by larger and mobile composite elements (all chimeric in nature since they result from an insertion of a transposon into a prophage) that are different, depending on whether the isolates are susceptible or resistant to tetracyclines [] (Varaldo, Montanari, & Giovanetti, 2009; Giovanetti, Brenciani, Lupidi, Roberts, & Varaldo, 2003; Banks, Porcella, Barbian, Martin, & Musser, 2003; Brenciani, et al., 2004; Giovanetti, Brenciani, Vecchi, Manzin, & Varaldo, 2005; Iannelli, Santagati, Oggioni, Stefani, & Pozzi, 2014). In tetracycline-susceptible isolates, a regular Tn1207.1 forms the left end of the 52-kb Φ1207.3 (formerly Tn1207.3) (Iannelli, Santagati, Oggioni, Stefani, & Pozzi, 2014; Santagati, et al., 2003), or is part of Φ10394.4 (ca. 59 kb) [] (Banks, Porcella, Barbian, Martin, & Musser, 2003; Banks, et al., 2004), with both integrated into the same chromosomal gene (comEC) and inserted into the same prophage (Varaldo, Montanari, & Giovanetti, 2009; Brenciani, et al., 2004). It appears that Φ1207.3 is more common than Φ10394.4 in mef(A)-positive, tetracycline-susceptible S. pyogenes isolates, and it has been shown that it was transferable both intra- and interspecifically (Varaldo, Montanari, & Giovanetti, 2009; Santagati, et al., 2003). In tetracycline-resistant isolates, evidence has emerged of a genetic linkage between mef(A) and tet(O) in a mobile phage-like element (Giovanetti, Brenciani, Lupidi, Roberts, & Varaldo, 2003; Giovanetti, Brenciani, Vecchi, Manzin, & Varaldo, 2005). Actually, this linkage corresponds to a variety of related tet(O)-mef(A) elements in which mef(A) is contained in a range of changeable and defective variants of Tn1207.1 (Brenciani, et al., 2004; Giovanetti, Brenciani, Vecchi, Manzin, & Varaldo, 2005). The most common representative is the transferable Φm46.1 element (ca. 60 kb) that is integrated into the chromosome within the 23S rRNA uracil methyltransferase gene (Giovanetti, Brenciani, Lupidi, Roberts, & Varaldo, 2003; Giovanetti, et al., 2014; Brenciani, Bacciaglia, Vignaroli, Pugnaloni, Varaldo, & Giovanetti, 2010). As described in S. pneumoniae, mef(E) is part of a typical mega element and is sometimes physically associated with tet(M) in the composite transposon Tn2009 [] (Del Grosso, et al., 2011). Note that both mef(A) and mef(E) alleles are always adjacent to the msr(D) gene originally named mel (Varaldo, Montanari, & Giovanetti, 2009; Ambrose, Nisbet, & Stephens, 2005). Similar to msr(A) in staphylococci, this gene also codes for an ABC family protein that contains the two prototypical ATP-binding domains, but lacks any obvious TMS. Consequently, it has not been clearly proven if this class 2 ABC protein functions as a drug exporter (Davidson, Dassa, Orelle, & Chen, 2008). Nonetheless, msr(D) alone is sufficient to confer a 64-, 128-, and 16-fold increase in MICs of erythromycin, clarithromycin, and telithromycin, respectively, but not to streptogramins, which distinguishes it from Msr(A) (Daly, Doktor, Flamm, & Shortridge, 2004).
mef(A)-carrying genetic elements (Varaldo, Montanari, & Giovanetti, 2009; Giovanetti E. , Brenciani, Lupidi, Roberts, & Varaldo, 2003; Brenciani, et al., 2004; Giovanetti E. , Brenciani, Vecchi, Manzin, & Varaldo, 2005; Iannelli, (more...)
Genetic elements carrying mef genes other than mef(A) (Varaldo, Montanari, & Giovanetti, 2009; Del Grosso, et al., 2011; Mingoia M. , et al., 2014; Mingoia M. , Morici, Brenciani, Giovanetti, & Varaldo, 2014). Open reading frames (ORFs) (more...)
Additional mef alleles have been detected in S. pyogenes, such as mef(I) and mef(O), as well as diverse mosaic variants (Sangvik, Littauer, Simonsen, Sundsfjord, & Dahl, 2005; Mingoia, et al., 2007). The subclass mef(I), 91% and 94% identical to mef(A) and mef(E), respectively, was first identified in S. pneumoniae and then among S. pyogenes clinical isolates (Blackman Northwood, et al., 2009; Mingoia, et al., 2007). In S. pneumoniae, mef(I) is embedded in a genetic element that also contains tet(M) and catQ. This element, designated 5216IQ complex (ca. 30 kb), consists of two portions, one derived from Tn5252 and Tn916 (which harbors a copy of tet(M) not expressed due to the lack of the promoter, the ribosome-binding site, and a part of the leader peptide) and another called IQ module, which encloses mef(I) and catQ, a gene that encodes a chloramphenicol acetyltransferase found in Clostridium perfringens [] (Mingoia, et al., 2007). In S. pyogenes, mef(I) and catQ are also linked through a partial fragment of the 5216IQ complex, designated the 5216IQ-like complex, with a defective IQ module and a partial or absent Tn916 (Del Grosso, et al., 2011). Note that all these IQ elements are ICEs that belong to the Tn5253 family, and are named ICESpy029IQ (ca. 55 kb) and ICESpy005IQ (ca. 50 kb) in S. pyogenes [] (Mingoia, et al., 2014; Mingoia, Morici, Brenciani, Giovanetti, & Varaldo, 2014). The mef(O) allele was first described in S. pyogenes, and exhibits 88% and 89% nucleotide identity with mef(A) and mef(E), respectively (Sangvik, Littauer, Simonsen, Sundsfjord, & Dahl, 2005).
Even if there are significant differences in the distribution of MLS resistance genes that depend on the country or year of isolation, the most common gene present in an international study appeared to be mef(A) (ca. 45%), followed by erm(B) (30%), and erm(TR) (ca. 25%) (Farrell, Morrissey, Bakker, & Felmingham, 2002).
Fluoroquinolones
Fluoroquinolones (FQs) are bactericidal agents that are widely used in both human and veterinary medicine. The targets of FQ molecules are the type II topoisomerases (namely, DNA gyrase and DNA topoismerase IV) that are both heterotetramers that consist of two subunits, GyrA2B2 and ParC2E2 (Hawkey, 2003). Due to the greater clinical use of FQ, bacterial resistance to these compounds has been increasingly reported during the last three decades. In Gram-positive cocci, FQ resistance results from target alterations due to point mutations that occur primarily in 120-bp conserved fragments, the so-called quinolone resistance-determining regions (QRDRs), of both parC and gyrA genes (Hooper, 2002). Active efflux of hydrophilic FQ molecules is also possible (Poole, 2005). Note that plasmid-mediated resistance has been recently described in Enterobacteriaceae, but has not yet been found in Gram-positive cocci (Cattoir & Nordmann, 2009).
In streptococci, high-level FQ resistance is only mediated by target modifications, whereas active efflux may confer low-level resistance in S. pneumoniae and viridans group streptococci (Guerin, Varon, Hoï, Gutmann, & Podglajen, 2000). However, the latter mechanism has not yet been detected in S. pyogenes (Malhotra-Kumar, et al., 2005). Importantly, the emergence and spread of FQ resistance may also be due to interspecies recombination within the parC QRDR and the acquisition of resistance via horizontal gene transfer from S. dysgalactiae subsp. equisimilis, which shares a global gene pool with S. pyogenes (Pletz, et al., 2006; Duesberg, et al., 2008; Pinho, Melo-Cristino, & Ramirez, 2010). Due to the lack of natural transformability of S. pyogenes, the genetic information is likely to be transferred by transduction through bacteriophages (Pinho, Melo-Cristino, & Ramirez, 2010). Different substitutions have been reported in clinical isolates of S. pyogenes: Ser81Ala/Tyr/Phe, Glu85Ala and Met99Leu in GyrA; Ser79Ala/Tyr/Phe, Asp83Asn, Asp91Asn, Ala121Val, Gly128Val, and Ser140Pro [] (Yan, et al., 2000; Alonso, Galimand, & Courvalin, 2002; Richter, et al., 2003; Reinert, Lütticken, & Al-Lahham, 2004; Albertí, et al., 2005; Orscheln, et al., 2005) Note that highly-resistant strains (levofloxacin MIC ≥16 mg/L) generally possess several mutations in the QRDRs of both gyrA and parC genes (Yan, Fox, Holland, Stock, Gill, & Fedorko, 2000; Richter, et al., 2003; Reinert, Lütticken, & Al-Lahham, 2004; Rivera, et al., 2005; Alonso, Mateo, Ezpeleta, & Cisterna, 2007; Wajima, et al., 2013), while low-level resistance (levofloxacin MIC, 2-4 mg/L) are commonly due to a single substitution in ParC (Alonso, Galimand, & Courvalin, 2002; Orscheln, et al., 2005; Rivera, et al., 2005; Wajima, et al., 2013; Yan, et al., 2008).
Amino acid sequences in S. pyogenes of quinolone resistance-determining regions (QRDRs) of GyrA (amino acids from 65 to 104, which correspond to positions 67-106 in E. coli numbering) and ParC (amino acids from 63 to 102, which correspond to positions (more...)
While high-level resistance remains exceptional among clinical isolates, FQ-non-susceptible strains that exhibit a low-level resistance (usually defined as ciprofloxacin MIC from 2 to 8 mg/L) may be common in some countries, such as Belgium (ca. 5% in 1999–2002, ca. 7% in 2003–2006), the USA (ca. 11% in 2002–2003), Spain (ca. 3% in 1999–2004, ca. 13% in 2005–2007), Portugal (ca. 5% in 1999–2006), Japan (ca. 16% in 2010–2012), and Italy (ca. 9% in 2012) (Malhotra-Kumar, et al., 2009; Malhotra-Kumar, et al., 2005; Orscheln, et al., 2005; Wajima, et al., 2013; Petrelli, et al., 2014). Notably, a vast majority of these isolates belong to emm-type 6, while some other genotypes (such as emm75 and emm89) have also been recorded (Malhotra-Kumar, et al., 2009; Malhotra-Kumar, Lammens, Chapelle, Mallentjer, Weyler, & Goossens, 2005; Montes, Tamayo, Orden, Larruskain, & Perez-Trallero, 2010; Yan, et al., 2008; Petrelli, Di Luca, Prenna, Bernaschi, Repetto, & Vitali, 2014; Alonso R. , Mateo, Galimand, Garaizar, Courvalin, & Cisterna, 2005). Indeed, emm-type 6 S. pyogenes seems to have an intrinsic reduced susceptibility to FQs, as a result of a polymorphism at position 79 of the parC gene (Orscheln, et al., 2005; Montes, Tamayo, Orden, Larruskain, & Perez-Trallero, 2010). However, despite this polymorphism, these strains are not more likely to develop high-level FQ resistance when compared to other emm types (Billal, et al., 2007).
Others
Glycopeptides
Glycopeptides (vancomycin and teicoplanin) are high-molecular weight molecules that interact with the D-Alanine-D-Alanine (D-Ala-D-Ala) termini of pentapeptide peptidoglycan precursors (Gold, 2001). They actually form a stable complex that involves five hydrogen bonds, which prevent the transglycosylation and transpeptidation reactions of the peptidoglycan synthesis.
In enterococci, resistance to glycopeptides is based on the presence of operons encoding enzymes i) that synthesize new precursors with low affinity where the last D-Ala residue is changed by a D-Lactate (D-Lac) or a D-Serine (D-Ser) residue; and ii) that eliminate or prevent the formation of a native precursor with a high affinity. Precursors with the D-Ala-D-Lac terminus have a 1,000-fold lower affinity to vancomycin than those ending in D-Ala-D-Ala, which results in a high-level resistance (MICs >16 µg/ml). On the other hand, the precursors that end in D-Ala-D-Ser have a 7-fold less affinity for vancomycin, which leads to a low-level resistance (MICs from 8 to 16 µg/ml) (Courvalin, 2006). Eight acquired operons (vanA, vanB, vanD, vanE, vanG, vanL, vanM, and vanN) and one intrinsic operon (vanC1-4) have been characterized to date (Watanabe, et al., 2009; Hegstad, Mikalsen, Coque, Werner, & Sundsfjord, 2010; Cattoir & Leclercq, 2013). The vanA, vanB, vanD, and vanM operons confer resistance through the synthesis of peptidoglycan precursors with C-terminal D-Ala-D-Lac residues, whereas isolates that harbor the vanC, vanE, vanG, vanL, and vanN operons contain precursors that end in D-Ala-D-Ser (Cattoir & Leclercq, 2013). Only vanA, vanB, and vanG have been identified in rare strains of Streptococcus spp. but never among S. pyogenes clinical isolates (Poyart, et al., 1997; Mevius, et al., 1998; Park, Nichols, & Schrag, 2014). The vanB gene cluster (vanB2 subtype) has only been identified in two Streptococcus lutetiensis isolates (MICs of vancomycin and teicoplanin at ≥256 and 3-4 µg/ml, respectively) as well as in two vanA-positive Streptococcus gallolyticus isolates (MICs of vancomycin and teicoplanin at ≥256 and 32-64 µg/ml, respectively) in France and the Netherlands (Poyart, et al., 1997; Mevius, et al., 1998; Dahl & Sundsfjord, 2003; Bjørkeng, Hjerde, Pedersen, Sundsfjord, & Hegstad, 2013). Two isolates of Streptococcus agalactiae and one of Streptococcus anginosus have been reported with low-level resistance to vancomycin (MIC of 4 µg/ml) in the US (Park, Nichols, & Schrag, 2014; Srinivasan, et al., 2014). Recently, a third strain of vanA-positive S. gallolyticus (MICs of vancomycin and teicoplanin both at ≥256 µg/ml) was described in Spain (Romero-Hernández, et al., 2015).
Sulfonamides and trimethoprim
Both sulfonamides and 2,4-diaminopyrimidines are synthetic agents and act as bacteriostatic inhibitors of bacterial biosynthesis of tetrahydrofolic acid, which is an essential cofactor for nucleic acid and protein syntheses (Masters, O'Bryan, Zurlo, Miller, & Joshi, 2003). They are generally given together due to a synergistic effect in vitro, and the most commonly used combination is sulfamethoxazole-trimethoprim, also called cotrimoxazole (or SXT) (Masters, O'Bryan, Zurlo, Miller, & Joshi, 2003). The sulfonamides are structural analogs of para-aminobenzoic acid that interfere with the formation of dihydropteroic acid, which is the first step in dihydrofolic acid synthesis, by blocking the enzymatic activity of the dihydropteroate synthase (DHPS). Trimethoprim inhibits a later step of the metabolic pathway (namely, the formation of tetrahydrofolic acid from dihydrofolic acid) by binding to the dihydrofolate reductase (DHFR) enzyme.
Bacterial resistance to sulfonamides results from chromosomal point mutations in the dhps (folP) gene coding for the natural DHPS, or the acquisition of plasmid-borne sul genes coding for resistant DHPS enzymes (Huovinen, Sundström, Swedberg, & Sköld, 1995). Resistance to trimethoprim in bacteria may be due to one or more of the following mechanisms: auxotrophy in thymine/thymidine; reduced intracellular antibiotic concentration (impaired permeability, active efflux); production of a naturally-insensitive DHFR; alterations and/or overexpression of the intrinsic DHFR encoded by the folA gene; or the presence of an additional plasmid-mediated dfr gene that codes for a resistant DHFR (Huovinen, Sundström, Swedberg, & Sköld, 1995).
Because of early inappropriate antimicrobial susceptibility testing (AST) approaches, S. pyogenes has been considered to be universally resistant to SXT for a long time. However, recent studies using standardized conditions (particularly AST media with low thymidine content) have confirmed the in vitro susceptibility of S. pyogenes to SXT (Bowen, et al., 2012). When appropriate methods are used, MICs of SXT are typically low [] and the prevalence of resistance among clinical isolates has ranged from 0% to 3% (Bowen, et al., 2012; Yourassowsky, Vanderlinden, & Schoutens, 1974; Eliopoulos & Wennersten, 1997). High-level sulfonamide resistance (MIC ≥128 µg/ml) in S. pyogenes is due to alterations within the chromosomally encoded DHPS (Swedberg, Ringertz, & Sköld, 1998; Jönsson, Ström, & Swedberg, 2003). Acquired resistance to trimethoprim in S. pyogenes may be due to the acquisition of transferable dfrF or dfrG genes (MICs from 32 to >512 µg/ml) or an amino acid substitution (Ile100Leu) in the intrinsic DHFR (MIC from 8 to 16 µg/ml) (Bergmann, Sagar, Nitsche-Schmitz, & Chhatwal, 2012; Bergmann, van der Linden, Chhatwal, & Nitsche-Schmitz, 2014).
Rifampin
Rifampin is a broad-spectrum antibiotic with bactericidal activity and is widely used for the treatment of tuberculosis. Like other members of the rifamycin family, it inhibits bacterial transcription through high-affinity binding to the DNA-dependent RNA polymerase (Lester, 1972). More precisely, rifampin interacts with the RNA polymerase β subunit encoded by the rpoB gene.
Rifampin resistance mainly results from chromosomal rpoB mutations that are responsible for target alterations, which occur at a frequency of ca. 10-6-10-8. These mutations are generally clustered in an 81-bp conserved fragment (the so-called rifampin resistance-determining region [RRDR]), which corresponds to codons 507-533 (according to Escherichia coli numbering) (Herrera, Jiménez, Valverde, García-Aranda, & Sáez-Nieto, 2003).
In S. pyogenes, rifampin resistance appears to be rare among clinical isolates (<0.5%) while MICs for wild-type strains are very low [] (Perez-Trallero, Urbieta, Montes, Ayestaran, & Marimon, 1998; Aubry-Damon, Galimand, Gerbaud, & Courvalin, 2002). High-level resistance to rifampin (>128 µg/ml) is due to RpoB changes at position 522 (Ser522Leu) (Aubry-Damon, Galimand, Gerbaud, & Courvalin, 2002; Herrera, et al., 2002).
Bacitracin
Like β-lactams and glycopeptides, bacitracin disrupts bacterial cell wall synthesis. It actually acts by preventing dephosphorylation and recycling of the lipid carrier (undecaprenol pyrophosphate) that allows the translocation of N-acetylmuramyl pentapeptide intermediates onto the surface of the cytoplasmic membrane (Butaye, Devriese, & Haesebrouck, 2003). Bacitracin is a narrow-spectrum antibiotic that is only active against Gram-positive bacteria.
Bacitracin is used in some topical preparations in human and veterinary medicine and has been employed in clinical laboratories as a presumptive marker for S. pyogenes identification. Indeed, among β-hemolytic streptococci, only S. pyogenes is susceptible to bacitracin. However, some bacitracin-resistant clones (MICs ≥16 µg/ml) have been uncommonly reported (York, Gibbs, Perdreau-Remington, & Brooks, 1999; Malhotra-Kumar, Wang, Lammens, Chapelle, & Goossens, 2003; Perez-Trallero, Garcia, Orden, Marimon, & Montes, 2004; Pires, et al., 2009). Notably, bacitracin resistance is commonly associated with macrolide resistance (cMLSB phenotype) mediated by the erm(B) gene with most of the isolates belonging to the emm28 genotype (Malhotra-Kumar, Wang, Lammens, Chapelle, & Goossens, 2003; Perez-Trallero, Garcia, Orden, Marimon, & Montes, 2004; Perez-Trallero, Garcia, Orden, Marimon, & Montes, 2004; Pires, et al., 2009; Mihaila-Amrouche, Bouvet, & Loubinoux, 2004). Even though the molecular mechanism of resistance is not well defined in this species, it might be associated with an overproduction of undecaprenol kinase encoded by the bacA gene, as shown in E. coli (Cain, Norton, Eubanks, Nick, & Allen, 1993; Chalker, et al., 2000). Notably, an ABC transporter (encoded by the bcrABD operon) responsible for bacitracin resistance in E. faecalis has not been detected among bacitracin-resistant S. pyogenes clinical isolates (Pires, et al., 2009; Manson, Keis, Smith, & Cook, 2004).
Chloramphenicol
The phenicols are inhibitors of bacterial protein synthesis and prevent peptide chain elongation after reversible binding to 23S rRNA (Schlünzen, et al., 2001). Chloramphenicol resistance is primarily due to the acquisition of chloramphenicol O-acetyltransferase (CAT) enzymes or to active efflux mediated by specific or multidrug transporters (Schwarz, Kehrenberg, Doublet, & Cloeckaert, 2004). Target modifications are also possible through point mutations or Cfr-mediated methylation in 23S rRNA (Schwarz, Kehrenberg, Doublet, & Cloeckaert, 2004; Kehrenberg, Schwarz, Jacobsen, Hansen, & Vester, 2005). CATs inactivate the antibiotic by acetylation in the three-step reaction, and there are two types of enzymes, according to their structure: type A CATs and type B CATs (also known as xenobiotic acetyltransferases or XATs) (Schwarz, Kehrenberg, Doublet, & Cloeckaert, 2004). Notably, chloramphenicol resistance is inducibly expressed through a translational attenuation regulation (Lovett, 1996).
Chloramphenicol resistance is rarely encountered in clinical isolates of β-hemolytic streptococci (ca. 0.5%), while it appears to be much more common among enterococcal species (ca. 40-50%) (Trieu-Cuot, et al., 1993). In S. pyogenes, only a few type-A CATs have been detected, such as cat(pC221) (group A-7), cat(pC194) (group A-9), catS (group A-12), and catQ (group A-16) (Del Grosso, et al., 2011; Schwarz, Kehrenberg, Doublet, & Cloeckaert, 2004; Trieu-Cuot, et al., 1993). Notably, mef(I) and catQ were demonstrated to be co-located in Tn5253-like ICEs (see above) [] and were cotransferred by transformation (Mingoia, et al., 2014).
Fusidic acid
Fusidic acid is a bacteriostatic antibiotic that inhibits bacterial protein synthesis by interfering with elongation factor G (EF-G) (Collignon & Turnidge, 1999). While it is very active against staphylococci (MIC50 and MIC90 at 0.12 and 0.25 µg/ml, respectively), β-hemolytic streptococci, including S. pyogenes, are less susceptible (MIC50 and MIC90 both at 4 µg/ml) [] (Jones, Mendes, Sader, & Castanheira, 2011). Acquired resistance has only been characterized in S. aureus, and is caused by mutations in the EF-G that encodes the fusA gene, or results from horizontal transfer of a plasmid-mediated determinant (fusB-E) (Turnidge & Collignon, 1999; O'Neill, McLaws, Kahlmeter, Henriksen, & Chopra, 2007). No S. pyogenes isolate highly resistant to fusidic acid has been reported to date.
Fosfomycin
Fosfomycin interferes with the cell wall synthesis at an early intra-cytoplasmic stage by specifically and irreversibly inhibiting the enzymatic activity of UDP-N-acetylglucosamine enolpyruvyltransferase (named MurA) (Falagas, Giannopoulou, Kokolakis, & Rafailidis, 2008). It exerts a slow bactericidal activity against a large panel of both Gram-positive and -negative bacteria, but is mainly used for staphylococcal infections and uncomplicated acute cystitis in young women (Patel, Balfour, & Bryson, 1997). Fosfomycin is moderately active against S. pyogenes with MICs from 2 to 64 µg/ml, while no high-level resistance has been reported in this species to date (Falagas, et al., 2010).