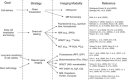
Direct labeling
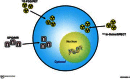
One of the most commonly used strategies for the labeling of stem cells for imaging in living subjects is that of direct labeling (Guzman et al., 2007; Thakur et al., 1976; Thakur et al., 1977). In a direct labeling strategy, labeling agents are introduced into the cells prior to transplantation (Figure ), stem cells transplanted and then followed in the living subject (Figure ). The strategy is then used to image the molecules previously introduced into the cell and use them as surrogate of the number of stem cells. Depending on the imaging modality to be used, cells can be labeled with quantum dots (Chakraborty et al., 2007; Lin et al., 2007; Rosen et al., 2007) or fluorophores (Askenasy et al., 2002; Keppler et al., 2006; Voura et al., 2004) for optical fluorescence imaging, superparamagnetic iron oxide particles (SPIO) for magnetic resonance imaging (MRI; Hill et al., 2003; Rogers et al., 2006), and radionuclides for single photon emission computed tomography (SPECT) or positron emission tomography (PET; Adonai et al., 2002; Bindslev et al., 2006; Kang et al., 2006; Rini et al., 2006).
Direct cell labeling. In a direct labeling strategy, labeling agents (for either MRI or radionuclide imaging) are introduced ex-vivo, stem cells are then transplanted to the tissue/organ of interest, and then non-invasive imaging is performed. SPIO: superparamagnetic (more...)
Imaging after direct cell labeling. In the left panel, cells have been loaded with paramagnetic particles (25 μg/mL medium), cells (2.8 to 16 × 107) delivered to the myocardium of an adult swine using fluoroscopy in a closed chest model, (more...)
Fluorescent semiconductor nanocrystals (known as quantum dots, QDs (Michalet et al., 2005) or different fluorophores (Kalchenko et al., 2006) have been used to monitor stem cells in the living subject. At each imaging time point, QDs and fluorophores are excited at predetermined wavelengths, and emit a fluorescent signal can be used for imaging of transplanted cells. This strategy has been used for tracking of stem cells in many organs (Kalchenko et al., 2006; Lin et al., 2007; Michalet et al., 2005; Seleverstov et al., 2006; Shah et al., 2007). However, fluorescence imaging has limited tissue penetration (around 2 mm) limiting the use of these techniques to superficial tissues in small animals (e.g., mice; Bengel et al., 2005; Massoud and Gambhir, 2003).
In this review, focus will be placed mainly on direct labeling modalities that have the potential to be applied clinically: MRI, SPECT, and PET.
Magnetic resonance imaging
Magnetic resonance imaging has been commonly used for imaging of stem cells, mostly based on concept of the imaging of super paramagnetic iron oxide (SPIOs). SPIOs are highly magnetic particles that can elicit changes in T2 relaxivity (effect known as T2*; Arbab et al., 2003; Bos et al., 2004; Di Tucci et al., 2008; Frank et al., 2003; Himes et al., 2004; Ittrich et al., 2007), allowing their detection in vivo (see Figure left). In a direct labeling strategy, SPIOs are incorporated into cells (using either electroporation or liposome-based incorporation techniques), cells then transplanted to the living subject, and subsequently MR imaging is performed (Bengel et al., 2005; Kraitchman et al., 2003) using gradient recalled echo sequences (Chen et al., 2008). The signal originated from the SPIOs is used as a surrogate for number of cells. MRI offers the advantage of high spatial resolution, resulting in detailed organ morphologic and functional information, and thus appears as a good candidate for an integrated stem cell imaging-functional assessment imaging approach in organs like the heart (Dick et al., 2003; Kraitchman et al., 2003) or the brain (Guzman et al., 2007; Watson et al., 2006). MR has also been used to monitor the delivery process. Specifically, MR fluoroscopy allows real-time assessment of the delivery of stem cells to the myocardium (Dick et al., 2003). However, the sensitivity of SPIO-based labeling is in the micromolar range (10−5 mol/L; Bengel et al., 2005; Massoud and Gambhir, 2003) and may not be sensitive enough to detect low signal levels (more than 1×105 cells are needed; Kraitchman et al., 2005). Sensitivity of the system can be increased using high-field magnets (e.g., 11 Tesla). However, for now these magnets are limited to use in small animal preclinical models. SPIO-based imaging constitutes a very good imaging strategy for initial localization of cells after transplantation, and for the co-registration of cell transplantation with areas of functional loss. After myocardial infarction, MR imaging is used to identify areas of infarction (using delayed enhancement MR techniques), and SPIO-MRI can be used to label stem cells for their localization immediately after delivery (Kraitchman et al., 2003; Figure left). However, SPIO-based imaging is not well suited for long-term monitoring of stem cells (Bengel et al., 2005; Chen et al., 2008; Li et al., 2008). Several reports suggested that SPIOs may not stay in the transplanted cells over time (Li et al., 2008), but rather be incorporated by macrophages and other cell types, and some iron may remain in the interstitial space. Because the effect of iron particles on the magnetic field continues (regardless of the location and status of the transplanted cells), there is an uncoupling between the MR signal and the viability of stem cells (Li et al., 2008). This effect may not be critical for the initial localization of transplanted cells, but will preclude the use of this strategy for the monitoring of transplanted stem cells over time.
Radionuclide imaging
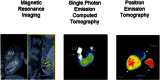
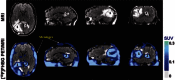
Radionuclide labeling of cells has also been used for cell imaging, using a strategy similar to SPIO-based techniques, which is to introduce a labeling agent to the cell prior to transplantation (see Figure ). Radionuclides used for this purpose can have different physical half lives (e.g., 99mTc: 6 hours, 111In: 2.8 days, 18F: 109 minutes, 64Cu: 12 hours), that will determine the amount of time that cells can be monitored non-invasively after cell labeling. For example, 111In-labeled cells have been used for many years to track the homing of inflammatory cells to localize inflammatory processes (Thakur et al., 1977; Thakur et al., 1977). More recently, the methodology has been applied to the labeling of stem cells, using different isotopes (e.g., 111In for SPECT, 18F-Fluoro-Deoxyglucose (18F-FDG) for PET). Use of isotopes like 18F-FDG (physical half life = 109 minutes) may allow tracking of cells for a 6–8 hours (after correction for isotope physical decay) after transplantation (Kang et al., 2006; Figure right), while use of 111In (see Figure middle) may allow cell tracking longer periods of time (up to 14 days; Chin et al., 2003). In addition to its physical half-life, each radionuclide also has a biological half-life (e.g., radionuclides may go in and out of the cell), which should be taken into consideration when performing these studies. One of the major advantages of SPECT and PET imaging is their high sensitivity (nano- and femto-molar, respectively), which permits the detection of relatively low amounts of signal (Adonai et al., 2002; Wu et al., 2004). However, SPECT and PET have relatively low spatial resolution, compared to other modalities (such as MRI), which may be a relative disadvantage for signal localization. The recent development of integrated PET-Computed Tomography (CT) and SPECT-CT provides a better anatomical guide for the location of the detected signal.
Advantages and disadvantages of direct labeling. Role in the assessment of stem cell therapy
Use of SPIOs (at least at doses of 20 pg of SPIO/cell) may have an effect on the gross morphology and proliferation capacity of stem cells, which may preclude their widespread use as a labeling strategy (Bos et al., 2004; Chen et al., 2008). In the case of radionuclide labeling, cell toxicity will likely vary depending on the radionuclide and dosage used (Carr et al., 1995; Zanzonico et al., 2006).
In addition, both SPIOs and radionuclides have a biological half-life (e.g., may go in and out of the cell), which should be taken into consideration when performing these studies. Thus, when using this imaging strategy, physical as well biological properties of the labeling agent should be considered together with the properties of the cells, to accurately determine the appropriate cell imaging modality. In general, the shortest half-life (biological or physical) will determine for how long transplanted cells can be monitored.
One of the main limitations of any direct labeling strategy is that it does not account for cell viability/division (i.e., cell numbers increase after cell division, but the number of radioisotope molecules stays the same), which results in “dilution” of the signal over time. This aspect of direct labeling limits its use for long term monitoring.
In summary, direct labeling strategies (whether it is MRI, SPECT-CT or PET-CT) appear to be a good imaging strategy for detection of cells shortly after transplantation (e.g., to ensure that cells where delivered to the intended organ or region of an organ), providing a good signal-to-noise ratio, but less suited for long-term monitoring of stem cell viability.
Reporter gene imaging
There is increasing need to understand the biology of stem cells after they are transplanted to the living subject. To achieve that, one needs a strategy that can accurately monitor stem cell biology in a longitudinal manner, without interfering with the normal biology of the cells under study or of the transplant recipient. In order to accurately study the biology of stem cells, such a system should be based on the physiologic activity of transplanted cells. Advances in molecular biology and imaging modalities have resulted in the development of reporter gene strategies, a system that allows evaluation of trans-gene expression in many disease states (see Figure ; Contag et al., 2000; Gambhir et al., 1999; Herschman, 2004; Inubushi and Tamaki, 2007; Kang and Chung, 2008; Massoud and Gambhir, 2003; Phelps, 2000; Wu et al., 2004; Zhang and Wu, 2007). Reporter genes consist of gene regulatory elements (promoters and enhancers) that drive the reporter gene DNA sequence, and a polyA sequence (which provides stabilization to the final product). Initially, reporter genes were mainly used in histology and ex-vivo studies (beta-galactosidase and luciferase) to assess the trans-gene expression in many different tissues and organs (Forss-Petter et al., 1990; Himes and Shannon, 2000; Naciff et al., 1999). Subsequently, reporter genes were used for imaging in vivo, using green fluorescent protein (GFP; Zhuo et al., 1997) or bioluminescence (Contag et al., 2000), providing one of the first evidences of the monitoring of trans-gene expression assessed in living subjects. For the application of stem cell monitoring, the reporter gene is incorporated into the cell before cell transplantation into the living subject. If the stem cells are viable (e.g., after transplantation), the reporter gene will be expressed and the protein (e.g., enzyme, cell surface receptor) will be encoded. On the other hand, if the reporter gene is not expressed due to cell death for example, no signal will be produced. At the specified imaging time point, an exogenously given substrate is administered. The interaction between the substrate and the encoded reporter protein (if present) will result in a signal, which can be detected non-invasively (using different imaging modalities, e.g. bioluminescence imaging, radionuclide imaging). The most common use of reporter genes is for the longitudinal study of stem cell viability. For this purpose, reporter genes are driven by a constitutive promoter (e.g., cytomegalovirus or CMV), which is always turned ON and, as long as the cell is viable and has the transcriptional machinery intact, will results in production of the reporter protein. However, the viral CMV promoter can undergo “gene silencing” (the gene is turned OFF) over time (Krishnan et al., 2006; a proposed mechanisms is linked to the large number of CpG repeats in the CMV promoter) that will result in reduced signal. From a practical standpoint, a decrease in the activity of the constitutive CMV promoter will result in decreased production of the reporter gene which could be mistaken for decreased stem cell survival. More recently, investigators have used promoters of constitutive mammalian cell proteins (e.g., β-actin, ubiquitin), which are proteins that are constantly produced in a cell. Most of these promoters undergo less gene silencing and stay “ON” as long as the cell is viable, and may represent better alternatives as the promoter of choice for assessment of stem cell viability. Furthermore, cells should be monitored for alterations in their phenotype and dividing capacity, although recent data suggest that the introduction of reporter genes do not seem to significantly alter the biological properties and differentiation capacity of stem cells (Cao et al., 2008; Wang et al., 2008; Wu et al., 2006).
Three different types of reporter gene/reporter probe strategies. (A) Enzyme-based bioluminescence imaging. D-Luciferin is a substrate molecular probe that is acted upon by the enzyme Firefly Luciferase to result in bioluminescence via a chemiluminescent (more...)
Specific biological pathways can also be investigated using reporter genes strategies. To study a certain pathway, a specific promoter is used (e.g., a protein specific promoter), and only when the intracellular signal for the production of that protein is active, would the reporter protein be made. For example, if one wants to monitor when embryonic stem cells differentiate into myocytes, one can use a reporter gene that is driven by a specific myocyte promoter, which will only be turned “ON” when the stem cell under study has turned “ON” the signaling cascade to produce a mature cardiac protein (e.g. troponin, desmin). Then, the activation of that specific promoter will “drive” the expression of the reporter gene, and one could visualize it non-invasively using different imaging modalities.
Fluorescent reporter genes
Use of GFP constitutes one of the first examples of the application of reporter genes for in vivo imaging of trans-gene expression. Fluorescence reporter proteins are very sensitive and result in the emission of a strong signal. However, both light excitation and emission undergo significant tissue attenuation (due to absorption) and tissue refraction, which limits the tissues that can be studied (up to 2 mm in depth; Contag, 2007; Massoud and Gambhir, 2003; Shah et al., 2004). In view of this, the most used application of fluorescent reporter genes in stem cell imaging is for ex-vivo analysis, where they can be used for cell sorting. Along these lines, the Gambhir laboratory and other groups have created a triple fusion reporter gene that has a fluorescent reporter gene, a bioluminescent reporter gene, and a PET reporter gene and that results in a fusion protein (Kesarwala et al., 2006; Ray et al., 2007). Cao et al. have transduced mouse embryonic stem cells (Cao et al., 2007; Cao et al., 2006), using the fluorescent protein (red fluorescent protein, mRFP) to identify the cells that have been effectively transduced with the fusion protein (using fluorescent activated cell sorting), and then transplanted those cells to the myocardium (Cao et al., 2007; Cao et al., 2006). Subsequently, the bioluminescent (firefly luciferase, Fluc) and PET reporter genes (herpes simplex virus enzyme thymidine kinase, HSV1-tk) were used for long-term monitoring of cell viability after transplantation (see next sections for details). Another common application of fluorescent reporter genes is for use in histology (immunofluorescence) for post-mortem confirmation of imaging results (Li et al., 2007). Using a similar approach to the one previously described (bi-or tri-fusion reporter genes), one can “mark” cells with a fluorescent reporter gene, and then when the tissue is excised, the “marked” cells can be easily identified using histological methods.
Bioluminescent reporter genes
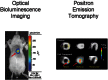
Reporter gene-bioluminescence imaging (BLI) is based on light emission and detection by specific cooled charge coupled device (CCD) cameras (Contag et al., 2000). Similar to other reporter gene strategies, the BLI signal is only emitted when cells are viable, and thus can be used for the longitudinal monitoring of stem cell survival and study of cell status (see Figure left). BLI is commonly used for the tracking of stem cells delivered to organs within small living animals (Chen et al., 2009; Rodriguez-Porcel et al., 2005; Wu et al., 2003). Fluc and Renilla luciferase (Rluc) are the two most common reporter genes used for BLI. BLI-Fluc imaging (see Figure left) is based on the oxidation of the substrate D-luciferin by the FLuc enzyme, a reaction that requires oxygen, magnesium, and ATP, and results in a red-shifted light emission (wavelength: 500–700nm). Imaging in the red-shifted light spectrum results in higher signal-to-background ratio, and thus makes it more “attractive” as a reporter gene for imaging in living subjects. On the other hand, Rluc does not require other cofactors and result in a lower wavelength emission (wavelength: 450– 550 nm), resulting in a lower signal-to-background ratio that makes it more challenging for imaging in the living subject. More recently, the Gambhir laboratory has developed red-shifted Rluc variants that have increased light output which can make Rluc a more attractive option for in vivo animal imaging (Loening et al., 2006; Loening et al., 2007).
Examples of reporter gene imaging. Examples of cell imaging using a reporter gene approach using the previously mentioned modalities. On the left panel, cells are genetically modified to carry the optical fluc reporter gene, delivered to the myocardium, (more...)
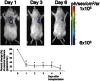
Using a reporter gene strategy, BLI has been successfully used for in vivo study of cell delivery and monitoring of stem cell viability in small living animals (see Figure ). BLI has been used for tracking and monitoring of different types of cells, such as neural cells (Okada et al., 2005), cardiomyoblasts (Chen et al., 2008; Rodriguez-Porcel et al., 2005; Wu et al., 2003) and embryonic stem cells (Cao et al., 2007; Cao et al., 2006). For the delivery of stem cells to the myocardium, investigators have used ultrasound guidance. This strategy (ultrasound guidance for the delivery of stem cells) allows the precise delivery of stem cells to the affected part of the organ (myocardium in this case; Rodriguez-Porcel et al., 2005; Springer et al., 2005). Furthermore, in these studies ultrasound guided delivery is complemented with non-invasive assessment of cardiac function (as it routinely done in the clinic). BLI has also been used to evaluate the response of stem cells transplanted to organs like the pancreas (Roth et al., 2006). Furthermore, using reporter genes, Kutscha et al have shown that modulation of the cellular and local microenvironment resulted in prolonged stem cell survival after transplantation to the myocardium (Kutschka et al., 2006), providing evidence that reporter gene imaging can be used to monitor changes in the biology of transplanted cells.
Longitudinal monitoring of stem cell survival using reporter gene imaging. Top, rat cardiomyoblasts were stably transfected with the CMV-fluc reporter gene, cells (2 × 106 cells), delivered to the myocardium of rats, and monitored using optical (more...)
As with other imaging modalities, BLI has some drawbacks and unsolved issues. Currently, BLI is mainly a planar imaging modality, providing limited depth information and signal localization within the living subject. Also, because of the lack of tomographic information, while BLI can provide information on the trafficking of stem cells to different organs, it may not be an ideal modality to accurately assess changes in stem cell viability as cells traffic between organs. This is because changes in the depth of the originated signal may be confused with changes in cell survival. Significant efforts are being devoted to the development of tomographic BLI either by rotating the subject under study, by detecting light using 2 or more cameras, or using spectral imaging (Lv et al., 2007; Soloviev, 2007). However, these systems are still under development. Due to the limited tissue depth that can be assessed with BLI, it is a technique that is mainly restricted to small animals (rats and mice), and very superficial tissues in larger living subjects (e.g., skin, in an intraoperative setting). In the future, these strategies may also be applied in specific clinical scenarios (e.g., skin, superficial tissues). Because BLI uses imaging strategies (reporter genes) similar to clinical modalities (e.g., PET and SPECT), BLI is routinely used as a starting step in the development of novel imaging strategies, and once the efficacy of these strategies is proven they can be adapted for use in the clinic.
In summary, BLI is a very useful imaging modality for the monitoring of stem cell trafficking and survival in small animals and will play a critical role in the study of cell biology and its interaction with their microenvironment.
Positron emission tomography/single photon emission computed tomography reporter genes
Both SPECT and PET reporter gene imaging are based on the interaction between an exogenously administered probe (that contains the tracer) and the reporter gene product (e.g., enzyme, receptor), what results in the retention of the radionuclide substrate that can then be imaged non-invasively (Bengel et al., 2005; Gambhir et al., 1998; Herschman, 2004; Tjuvajev et al., 1998; Wu et al., 2004). One of the main advantages of PET and SPECT imaging is that it can provide tomographic, quantitative, and volumetric information, allowing one to better localize and quantify the detected signal within the subject under study (see Figure right). In addition, the sensitivity of PET is in the femtomolar range (10−12 mol/L), higher than MR imaging (10−5 mol/L), but not as sensitive as optical imaging (at limited depths, 10−15 mol/L). There are predominantly three reporter gene systems (for PET or SPECT) that have been used for cell imaging. The system used the most is based on the production of an intracellular enzyme (e.g., HSV1-tk), that phosphorylates an exogenously administered substrate which is retained in the cell due to its negative charge. While normal cells (without the HSV1-tk) do carry the enzyme mammalian wild type thymidine kinase, it only minimally phosphorylates the radionuclide probes used in this system. On the other hand, in cells carrying the HSV1-tk, the exogenously administered probe undergoes significant phosphorylation and intracellular retention, leading to a robust signal-to-background ratio, enabling accurate monitoring of these cells. Furthermore, this strategy is very powerful because the enzyme can phosphorylate many molecules of the radionuclide substrate, increasing the signal retained in the cells of interest and improving the signal-to-background ratio. However, the probe has to cross the cell membrane, which may limit the interaction between substrate and enzyme, resulting in reduced signal. In addition, because it is a non-mammalian protein (of viral origin) it has the potential to trigger an immunological response, resulting in a reaction in the organism with decreased overall signal. Attempts to circumvent this problem have included use of a mammalian protein (dopamine receptor, see later in this paragraph) or a mammalian mitochondrial tk (Ponomarev et al., 2007), or a destabilized HSV1-tk (Hsieh et al., 2008). A second reporter gene imaging strategy is based on the imaging of dopamine receptor using PET (D2R, MacLaren et al., 1999). In this case, the reporter gene encodes for a cell membrane protein, which binds to an exogenously given probe, and the “bound probe” is then imaged non-invasively with PET. It is important to mention that the wild type D2R has the potential to elicit a downstream biological response, which can be prevented if one uses a mutant version of the dopamine receptor (Chen et al., 2004; Liang et al., 2001). Importantly, the mutant version of the D2R (Mutation of Asp80 or Ser194) maintains the affinity for the PET probe (3-(2’-[18F]-fluoroethyl)-spiperone) used. This strategy can be advantageous as the probe does not have to cross the cell membrane in order to interact with the reporter protein. However, this approach is limited by the amount of signal that can be produced, as one receptor interacts with only one molecule of the ligand. One of the most common strategies is the use of a PET-based reporter gene strategy for the visualization of transplanted cells to the myocardium and showed that it can be used to monitor the fate of stem cells after transplantation to the living subject (Li et al., 2007; Wu et al., 2003). Similarly, we have shown that transplantation of pancreatic islet cells can be followed non-invasively (Kim et al., 2006; Lu et al., 2006; Lu et al., 2006). A third approach consists on the encoding of the sodium-iodide symporter (NIS; Kang et al., 2005; Kim et al., 2005; Miyagawa et al., 2005; Terrovitis et al., 2008), which is a thyroid transmembrane protein, under physiological conditions transport iodine into the cells, in exchange for sodium. Successful sequencing and cloning of the NIS gene sequence allowed its use as a reporter gene in the thyroid and other organs. This system has been used not only for PET (with 124I as the tracer), but also for SPECT imaging (using 123I or 99Tc-perthechnetate as tracer). The somatostatin receptor reporter gene (Zinn and Chaudhuri, 2002) can also be evaluated with both SPECT and PET as imaging modalities, providing flexibility on the assessment of trans-gene expression (Schillaci, 2007; Sharma et al., 2002; Zhernosekov et al., 2005). Other less frequent reporter genes include the neurotensin receptor subtypes (Alvarez-Maya et al., 2001; Tavares et al., 1999), and cytosine deaminase (Kreuzer et al., 1996; Lee et al., 2007). Details of the use of these reporter genes are beyond the scope of this review and can be found elsewhere (Sharma et al., 2002).
The major advantage of PET and SPECT (when compared to BLI) resides in their potential for clinical use. Both PET and SPECT radionuclides are of relatively high energy (PET: 511KeV, SPECT: 80–250 KeV) and do not undergo significant tissue attenuation. While SPECT (due to its relatively low energy) does have some tissue attenuation, it does not preclude its use in patients and proof of that is its extensive clinical use.
These imaging modalities present a number of issues that need to be considered from the operational standpoint. On one side, PET has significant flexibility for the production of specific probes for the detection of different processes in the living subject (almost any compound can be labeled with a radionuclide), which is a significant advantage as it allows the researcher to first identify the molecule that needs to be studied, and then design a specific probe that will target that molecule. However, the production of PET probes is complex, needs advanced chemistry and very tight quality control. In addition, depending on the half-life of the radioisotope used, it requires an on-site (or at least near-by) cyclotron, that limits this strategy to medium to large research centers. From the imaging standpoint, all electron-positron annihilations (whether is from 18F, 64Cu or 11C) result in the production of photons of 511KeV, and as such we can not detect differences in registered signals. SPECT, on the other hand, can detect simultaneous signals of different energies (by varying the detection windows, as it is routinely done with the perfusion agents 201Tl and 99Tc). At the same time, tracer labeling is less complex (compared to PET) and, for the most part, can be done in a radionuclide pharmacy. However, the spatial resolution of SPECT is less than that of PET, and this variable may be of importance when we try to spatially localize relatively low number of cells. From the tracer perspective, SPECT labeling can be somewhat limited by its chemistry, with the result that not every compound can be easily labeled, thus providing less labeling flexibility compared to PET. In practical terms, if the compound of interest can be labeled with SPECT and answers the research questions posed by the investigator, its production and availability is more accessible to many academic and research centers.
Magnetic Resonance reporter genes
Over the last few years, significant efforts have been devoted to combined these modalities (MRI and reporter gene technology) and develop MR reporter genes (Cohen et al., 2005; Gilad et al., 2007).
MRI-based reporter genes efforts are based on the production of proteins, mostly intracellular metalloproteins (transferring, ferritin, tyrosinase; Gilad et al., 2007). As previously described, iron is a paramagnetic substance that induces changes in relaxivity (i.e., T2* effect) that can be detected using specific imaging sequences. Physiologically, iron enters cells through the transferrin receptor (TfR) that binds the transferrin protein containing two iron atoms and internalizes iron molecules. So the goal of this strategy is to express transferring, that will accumulate large quantities of iron intracellularly for non-invasive detection.
A reporter gene strategy targeted to express Ferritin has also been used for MR-based detection. Ferritin is a metalloprotein that functions as the body's iron depot and can contain up to 4000 iron atoms. Native ferritin is in essence an anti-magnetic particle, but several orders of magnitude weaker than SPIOs. Considerable efforts are being devoted to improve its relaxivity by removing its native core (oxyhydroxide) and reconstituting the protein shell with a superparamagnetic core.
Tyrosinase has also been used as a MR reporter gene (Weissleder et al., 1997). Briefly, tyrosinase participates in the production of melanin, and melanin has high affinity for iron, leading to increased relaxivity. Tyrosinase has been transfected to fibroblasts and embryomal kidney cells as well as breast cancer cells which resulted in increased signals. However, there is concern regarding potential toxicity of iron. Melanin production produces reactive oxygen species (an important and deleterious component of the oxidative stress cascade) and thus can exhibit significant toxic effects.
There are a few drawbacks in the use of metalloproteins as MR reporter genes that also deserve consideration (Gilad et al., 2007). First, the signal is dependent on the accumulation of iron inside the cells and for how long iron particles can be retained inside cells, what may result in toxicity to the cell. Second, when cells divide the signal gets diluted and the “clock” starts again, as cells need to start again to accumulate enough iron for MRI detection, making it difficult to interpret correlation between detected signal and viability of transplanted cells. From the imaging standpoint, the relaxation is dynamic and dependent on the iron loading conditions. R2 relaxivity is high at low iron doses, it decreases at intermediate iron loading conditions and when iron conditions are high, T2 relaxation remains constant, which may preclude accurate quantification of the obtained signal. As previously described for direct cell labeling, MR signal works on the basis of accumulation of iron. Similar to what happens with SPIOs, when cells die the accumulated iron continues to be present inside the cells for some time (until cells are dissolved or phagocytosed by macrophages). The remaining iron continues to induce a change in MR relaxivity (even if cells are not alive), and thus the signal is not representative nor linear of cell viability (Li et al., 2008).