

 .
.Bookshelf ID: NBK3
NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.

The Human ATP-Binding Cassette (ABC) Transporter Superfamily
Michael Dean.
Author Information and AffiliationsBethesda (MD): National Center for Biotechnology Information (US); 2002 Nov 18.
Excerpt
The ATP-binding cassette (ABC) transporter superfamily contains membrane proteins that translocate a wide variety of substrates across extra- and intracellular membranes, including metabolic products, lipids and sterols, and drugs. Overexpression of certain ABC transporters occurs in cancer cell lines and tumors that are multidrug resistant. Genetic variation in these genes is the cause or contributor to a wide variety of human disorders with Mendelian and complex inheritance including cystic fibrosis, neurological disease, retinal degeneration, cholesterol and bile transport defects, anemia, and drug response phenotypes. Conservation of the ATP-binding domains of these genes has allowed the identification of new members of the superfamily based on nucleotide and protein sequence homology. Phylogenetic analysis places the 48 known human ABC transporters into seven distinct subfamilies of proteins. For each gene, the precise map location on human chromosomes, expression data, and localization within the superfamily have been determined. These data allow predictions to be made as to potential function(s) or disease phenotype(s) associated with each protein. Comparison of the human ABC superfamily to that of other sequenced eukaryotes including Drosophila indicated that there is a rapid rate of birth and death of ABC genes and that most members carry out highly specific functions that are not conserved across distantly related phyla.
Introduction to ABC Protein and Gene Organization
The ATP-binding cassette (ABC) genes represent the largest family of transmembrane (TM) proteins. These proteins bind ATP and use the energy to drive the transport of various molecules across all cell membranes (1–3) (Figure 1). Proteins are classified as ABC transporters based on the sequence and organization of their ATP-binding domain(s), also known as nucleotide-binding folds (NBFs). The NBFs contain characteristic motifs (Walker A and B), separated by approximately 90–120 amino acids, found in all ATP-binding proteins (Figure 1). ABC genes also contain an additional element, the signature (C) motif, located just upstream of the Walker B site (4). The functional protein typically contains two NBFs and two TM domains (Figure 2). The TM domains contain 6–11 membrane-spanning α-helices and provide the specificity for the substrate. The NBFs are located in the cytoplasm and transfer the energy to transport the substrate across the membrane. ABC pumps are mostly unidirectional. In bacteria, they are predominantly involved in the import of essential compounds that cannot be obtained by diffusion (sugars, vitamins, metal ions, etc.) into the cell. In eukaryotes, most ABC genes move compounds from the cytoplasm to the outside of the cell or into an intracellular compartment [endoplasmic reticulum (ER), mitochondria, peroxisome]. Most of the known functions of eukaryotic ABC transporters involve the shuttling of hydrophobic compounds either within the cell as part of a metabolic process or outside the cell for transport to other organs, or for secretion from the body.


Figure 2
ABC gene structure. A diagram of an ABC half transporter and a full transporter. The half transporter can form homo- or heterodimers, whereas the entire full transporter is found in one transcript.
The eukaryotic ABC genes are organized either as full transporters containing two TMs and two NBFs, or as half transporters (4) (Figure 2). The latter must form either homodimers or heterodimers to form a functional transporter. ABC genes are widely dispersed in eukaryotic genomes and are highly conserved between species, indicating that most of these genes have existed since the beginning of eukaryotic evolution. The genes can be divided into subfamilies based on similarity in gene structure (half versus full transporters), order of the domains, and on sequence homology in the NBF and TM domains. There are seven mammalian ABC gene subfamilies, five of which are found in the Saccharomyces cerevisiae genome (5).
A list of Web resources on ABC genes and products can be found in Box 1.
Box 1
ABC transporter superfamily web resources.
A more detailed account of each of the human ABC genes is given below. For each gene, a concise description is given on the known function and disease involvement, and links to other databases, such as UniGene, OMIM, and GenBank, are given where appropriate. This is a comprehensive treatment: even genes that are very poorly characterized are included. For genes such as CFTR and ABCB1 /PGP/MDR that have been studied extensively, a brief review is given with links to other resources and review articles. Suggested corrections and additions are welcome for future updates of these pages and should be sent to the author (vog.frcficn@naed).
Nomenclature
All human and mouse ABC genes have standard nomenclature, developed by the Human Genome Organization (HUGO) at a meeting of ABC gene researchers. Details of the nomenclature scheme can be found at: http://www.genenames.org/genefamily/abc.html.
Researchers working on ABCC7 /CFTR, ABCB2 /TAP1, and ABCB3 /TAP2 have petitioned to keep their original gene designations. Official gene symbols are used in this monograph, but all known synonyms are also included to allow researchers to refer to the literature.
Overview of Human ABC Gene Subfamilies
A list of all known human ABC genes is displayed in Table 1. This list includes an analysis of the released genome sequences (6, 7). An analysis of the genome sequence indicates the presence of at least 19 pseudogenes (Dean, unpublished). There remain several sequences in the genome with homology to ABC genes that lie in incompletely sequenced regions and may represent additional pseudogenes or functional loci.
Table 1
List of human ABC genes, chromosomal location, and function.
By aligning the amino acid sequences of the NBF domains and performing phylogenetic analysis with a number of methods, the existing eukaryotic genes can be grouped into seven major subfamilies. A few genes do not fit into these subfamilies, and several of the subfamilies can be further divided into subgroups.
ABCA (ABC1)
The human ABCA subfamily comprises 12 full transporters (Table 1) that are further divided into two subgroups based on phylogenetic analysis and intron structure (8, 9). The first group includes seven genes dispersed on six different chromosomes ( ABCA1 , ABCA2 , ABCA3 , ABCA4 , ABCA7 , ABCA12 , ABCA13 ), whereas the second group contains five genes ( ABCA5 , ABCA6 , ABCA8 , ABCA9 , ABCA10 ) arranged in a cluster on chromosome 17q24. The ABCA subfamily contains some of the largest ABC genes, several of which are over 2,100 amino acids long. Two members of this subfamily, the ABCA1 and ABCA4 (ABCR) proteins, have been studied extensively. The ABCA1 protein is involved in disorders of cholesterol transport and HDL biosynthesis (see below). The ABCA4 protein transports vitamin A derivatives in the outer segments of rod photoreceptor cells and therefore performs a crucial step in the vision cycle.
The ABCA genes are not present in yeast; however, evolutionary studies of ABCA genes indicate that they arose as half transporters that subsequently duplicated, and that certain sets of ABCA genes were lost in different eukaryotic lineages (10).
ABCB (MDR/TAP)
The ABCB subfamily is unique in mammals in that it contains both full transporters and half transporters. Four full transporters and seven half transporters have currently been described as members of this subfamily. ABCB1 (MDR/PGY1) is the first human ABC transporter cloned and characterized through its ability to confer a MDR phenotype to cancer cells. The physiological functional sites of ABCB1 include the blood-brain barrier and the liver. The ABCB4 and ABCB11 proteins are both located in the liver and are involved in the secretion of bile acids. The ABCB2 and ABCB3 (TAP) genes are half transporters that form a heterodimer to transport peptides into the ER that are presented as antigens by the class I HLA molecules. The closest homolog of the TAPs, the ABCB9 half transporter, has been localized to lysosomes. The remaining four half transporters, ABCB6, ABCB7, ABCB8, and ABCB10, localize to the mitochondria, where they function in iron metabolism and transport of Fe/S protein precursors.
ABCC (CFTR/MRP)
The ABCC subfamily contains 12 full transporters with a diverse functional spectrum that includes ion transport, cell-surface receptor, and toxin secretion activities. The CFTR protein is a chloride ion channel that plays a role in all exocrine secretions; mutations in CFTR cause cystic fibrosis (11). ABCC8 and ABCC9 proteins bind sulfonylurea and regulate potassium channels involved in modulating insulin secretion. The rest of the subfamily is composed of nine MRP-related genes. Of these, ABCC1, ABCC2, and ABCC3 transport drug conjugates to glutathionine and other organic anions. The ABCC4, ABCC5, ABCC11, and ABCC12 proteins are smaller than the other MRP1-like gene products and lack an N-terminal domain (12) that is not essential for transport function (13). The ABCC4 and ABCC5 proteins confer resistance to nucleosides including PMEA and purine analogs. The human genome contains a seemingly intact ABCC gene on chromosome 21 (ABCCxP1) that contains a frameshift in one exon and is therefore a pseudogene. The same frameshift mutation is present in the gorilla and chimpanzee homologs, but the gene appears to be functional and expressed in monkeys (Annilo et al., in preparation).
ABCD (ALD)
The ABCD subfamily contains four genes in the human genome and two each in the Drosophila melanogaster and yeast genomes. The yeast PXA1 and PXA2 products dimerize to form a functional transporter involved in very long chain fatty acid oxidation in the peroxisome (14). All of the genes encode half transporters that are located in the peroxisome, where they function as homo- and/or heterodimers in the regulation of very long chain fatty acid transport.
ABCE (OABP) and ABCF (GCN20)
The ABCE and ABCF subfamilies contain gene products that have ATP-binding domains that are clearly derived from ABC transporters but they have no TM domain and are not known to be involved in any membrane transport functions. The ABCE subfamily is solely composed of the oligo-adenylate-binding protein, a molecule that recognizes oligo-adenylate and is produced in response to infection by certain viruses. This gene is found in multicellular eukaryotes but not in yeast, suggesting that it is part of innate immunity. Each ABCF gene contains a pair of NBFs. The best-characterized member, the S. cerevisiae GCN20 gene product, mediates the activation of the eIF-2α kinase (15), and a human homolog, ABCF1, is associated with the ribosome and appears to play a similar role (16).
ABCG (White)
The human ABCG subfamily is composed of six “reverse” half transporters that have an NBF at the N terminus and a TM domain at the C terminus. The most intensively studied ABCG gene is the white locus of Drosophila. The white protein, along with brown and scarlet, transports precursors of eye pigments (guanine and tryptophan) in the eye cells of the fly (17). The mammalian ABCG1 protein is involved in cholesterol transport regulation (18). Other ABCG genes include ABCG2 , a drug-resistance gene; ABCG5 and ABCG8 , coding for transporters of sterols in the intestine and liver; ABCG3 , to date exclusively found in rodents; and the ABCG4 gene that is expressed predominantly in the liver. The functions of the last two genes are unknown.
ABC Genes and Human Genetic Disease
Many ABC genes were originally discovered during the positional cloning of human genetic disease genes. To date, 14 ABC genes have been linked to disorders displaying Mendelian inheritance (19) (Table 2). As expected from the diverse functional roles of ABC genes, the genetic deficiencies that they cause also vary widely. Because ABC genes typically encode structural proteins, all of the disorders are recessive or X-linked recessive and are attributable to a severe reduction or lack of function of the protein. However, heterozygous variants in ABC gene mutations are being implicated in the susceptibility to specific complex disorders.
Table 2
Diseases and phenotypes caused by ABC genes.
Few ABC gene mutations are lethal. Untreated cystic fibrosis ( ABCC7 /CFTR) is typically lethal in the first decade, and adrenoleukodystrophy ( ABCD1 /ALD) can also be fatal in the first 10 years of life. The only mutations described in ABCB7 are missense alleles, and the yeast homolog is essential to mitochondria, suggesting that this gene is essential. The only developmental defect ascribed to an ABC gene is the congenital absence of the vas deferens that occurs in both cystic fibrosis patients and patients with less severe alleles that present male sterility as their only phenotype. Thus, most ABC genes do not play an essential role in development.
Mouse Knockouts
Most of the human genes have a clear mouse ortholog; however, there are several exceptions (Table 3). Several ABC genes have been disrupted in the mouse (Table 3). These include some of the genes mutated in human diseases, as well as several of the known drug transporters. The Abca1 and Cftr –/– mice show compromised viability; however, the remaining knockouts are viable and fertile, and many show either no phenotype or a phenotype only under stressed conditions.
Table 3
ABC genes: human and mouse orthologs.
Multidrug Resistance and Cancer Therapy
Cells exposed to toxic compounds can develop resistance by a number of mechanisms including decreased uptake, increased detoxification, alteration of target proteins, or increased excretion. Several of these pathways can lead to multidrug resistance (MDR) in which the cell is resistant to several drugs in addition to the initial compound. This is a particular limitation to cancer chemotherapy, and the MDR cell often displays other properties, such as genome instability and loss of checkpoint control, that complicate further therapy. ABC genes play an important role in MDR, and at least six genes are associated with drug transport.
Three ABC genes appear to account for nearly all of the MDR tumor cells in both human and rodent cells. These are ABCB1 /PGP/MDR1, ABCC1 /MRP1, and ABCG2 /MXR/BCRP (Table 4). No other genes have been found overexpressed in cells that display resistance to a wide variety of drugs and in cells from mice with disrupted Abcb1a , Abcb1b , and Abcc1 genes; the Abcg2 gene was overexpressed in all MDR cell lines derived from a variety of selections (20).
Table 4
ABC transporters involved in drug resistance.
Inhibitors of the major ABC genes contributing to MDR have been developed, and extensive experimentation and clinical research have been performed to attempt to block the development of drug resistance during chemotherapy (Table 4). The latest experiments with high-affinity and high-specificity ABCB1 inhibitors show that the gene is expressed in many primary tumors in human patients and that its activity can be blocked with doses of inhibitor that do not have adverse side effects or disrupt the pharmacology of the drug regimen (21). Thus, the development of highly specific inhibitors to the other major drug transporters could lead to the development of much more effective chemotherapy protocols.
Another limitation of chemotherapy is the narrow difference in sensitivity of the tumor cells to drugs and sensitivity of the patient's normal stem cells. ABC genes have also been used as tools to deliver drug transporters to early stem cells and to protect them from chemotherapeutic drugs. This strategy would allow high doses of drug to be given for longer periods of time.
Phylogenetic Analysis of Human ABC Genes
The identification of the complete set of human ABC genes allows a comprehensive phylogenetic analysis of the superfamily. Alignment of the NBFs from each gene and a neighbor-joining tree resulting from this analysis is displayed (Figure 3). The subclassification of ABC transporters is in excellent agreement with the phylogenetic trees obtained. In particular, all major ABC transporter families are represented in the human tree by stable clusters with high statistical significance.

This analysis provides compelling evidence for frequent domain duplication of ATP-binding domains in ABC transporters. Virtually invariably, both ATP-binding domains within a gene are more closely related to each other than to ATP-binding domains from ABC transporter genes of other subfamilies. This could represent a concerted evolution of domains within the same gene, but this seems unlikely because the two domains within each gene are substantially diverged. Therefore, it appears that duplication of ATP-binding domains within major ABC families was a result of several independent duplication events rather than a single ancestral duplication.
Mouse ABC Genes
Analysis of the Celera assembly of the mouse genome was used to identify homologs of the human ABC genes. With only a few exceptions, there is concordance between the two mammalian species (Table 3). The exceptions are a duplicated copy of the ABCB1 /PGP/MDR gene ( Mdr1b ), an ABCG family gene related to ABCG2 that is present in the mouse and not in the human ( Abcg3 ) (22), loss of Abcc11 (Dean, unpublished), duplication of the ABCA8 gene in the mouse ( Abca8a ), and a loss in the mouse of ABCA10 (Annilo et al., submitted). In addition, mice have a cluster of three ABCA family genes that is not characterized in the human genome (Chen, Annilo, Shulenin, and Dean, unpublished). This region of the human genome is incompletely characterized and does not currently contain any described functional loci. Therefore, mice have 52 ABC genes and most of the human genes have a single homolog in the mouse genome, indicating that the functions of the mouse genes should be highly similar to human genes.
Drosophila ABC Genes
The organization and annotation of the Drosophila ABC genes have been determined from the Celera (23) and Flybase (5) databases. Initial subfamily classifications were assigned based on homology and BLAST scores, and the location of each gene is shown (Table 5). In total, there are 56 genes with at least one representative of each of the known mammalian subfamilies (Table 6). The subfamily groupings were confirmed by phylogenetic analyses. A representative tree is shown in Figure 4. As expected, genes from the same subfamily cluster together and confirm the initial assignments made by inspection.
Table 5
Drosophila ABC genes.
Table 6
ABC gene subfamilies in characterized eukaryotes.

Figure 4
Phylogenetic tree of the human and Drosophila G subfamily ABC genes. An alignment of the G family genes from Drosophila and human genomes were aligned. Analysis was performed as described for Figure 3 (Annilo and Dean, unpublished).
As in the human and yeast genomes, the Drosophila ABC genes are largely dispersed in the genome. There are four clusters of two genes and one cluster of four genes (Figure 5). One of these clusters (on chromosome 2L, band 37B9) is composed of an ABCB and an ABCC gene, indicating that this is a chance grouping of genes. The remaining clusters are composed of genes from the same subfamily and are arranged in a head-to-tail fashion, consistent with gene duplication. Because the clusters are themselves dispersed and involve different subfamilies, they presumably represent independent gene duplication events.

Figure 5
Map of the Drosophila ABC genes. A diagram of each Drosophila chromosome is shown with the location and gene subfamily designation of each gene.
The best-studied Drosophila ABC genes are the eye pigment precursor transporters white ( w ), scarlet ( st ), and brown ( bw ). These genes are part of the ABCG subfamily and have a unique NBF-TM organization. Surprisingly, there are 15 ABCG genes in the fly genome, making this the most abundant ABC subfamily. This is in sharp contrast to the five or six known ABCG genes in the human and mouse genomes, respectively. The Drosophila ABCG genes are highly dispersed in the genome with only two pairs of linked genes. In addition, they are very divergent phylogenetically, suggesting that there were many independent and ancient gene duplication events. The Atet gene is the only Drosophila ABCG family gene that has a close ortholog in the human genome ( ABCG1 and ABCG4 ) (Figure 4).
Several Drosophila ABCB genes, Mdr49 , Mdr50 , and Mdr65 , have also been well characterized. A fourth member of this group, CG10226 , found clustered with Mdr65 , was also identified (Table 5). These genes are closely related to the human and mouse P-glycoproteins ( ABCB1 and ABCB4), and disruption of Mdr49 results in sensitivity to colchicines (24).
Phenotypic mutants that are not assigned to genes and lie in the region of Drosophila ABC genes are shown (Table 5). The most promising connection is the identification of several eye phenotypes (vin, rose, cln) in the region of the CG7346 gene. Because CG7346 is part of the ABCG family and is therefore related to w , st , and bw , it is tempting to speculate that mutations in CG7346 cause one or more of these phenotypes. Because ABC genes perform very diverse functions and are associated with varied phenotypes, it is hard to gather much additional insight from this analysis.
Three genes, CG9990 , CG6162 , and CG11147 , were identified that do not fit into any of the known subfamilies and, in fact, are most closely related to ABC genes from bacteria. These genes are within large contigs and have introns and therefore do not represent contamination from bacterial sequences. This group forms a distinct cluster on the Drosophila tree. This new ABC transporter subfamily in Drosophila is significantly different from all known families of ABC transporters and might play an as-yet-unidentified functional role. These genes have been designated as subfamily H.
Because of the high rate of birth and death of ABC genes, very few Drosophila genes have a human ortholog. This indicates that the genes have evolved to carry out functions that are specialized to insects and mammals. This is borne out by the experimental data to date. For example, the insect and vertebrate eyes are convergent organs, and the eye pigment transporters in flies have no comparable functional homolog in vertebrates. Similarly, the vertebrate ABCA4 (photoreceptor-specific transporter), CFTR (chloride channel controlling exocrine secretion), and most other mammalian ABC genes have specialized functions that are not present in insects and nematodes. Therefore, the genetic and functional analysis of Drosophila genes is not likely to lead to the direct understanding of the function of the individual mammalian ABC genes.
ABCA Genes
ABCA1
The ABCA1 gene was identified in the mouse and human genomes and mapped to human chromsome 9q31 and mouse chromosome 4, 23.1 cM (25). It was subsequently found that ABCA1 is the causative gene in Tangier disease, a disorder of cholesterol transport between tissues and the liver, mediated by binding of the cholesterol onto high-density lipoprotein (HDL) particles (26–30). Patients with familial hypoalphalipoproteinemia have also been described that have mutations in the ABCA1 gene, demonstrating that these disorders are allelic (31). Other patients with reduced levels of HDLs without the classical symptoms of Tangier disease have also been described with ABCA1 mutations (32).
ABCA1 controls the extrusion of membrane phospholipid and cholesterol toward specific extracellular acceptors; however, the exact role of the protein in this process is not known. It has been proposed that ABCA1 carries out the flipping of membrane phospholipid, principally phosphatidylcholine, toward the lipid-poor, nascent apolipoprotein particle, which can now accept cholesterol (33). The ABCA1-dependent control on the lipid content of the membrane dramatically influences the plasticity and fluidity of the membrane itself and, as a result, affects the lateral mobility of membrane proteins and/or their association with membrane domains of special lipid composition. ABCA1 also plays a role in the engulfment of apoptotic bodies. Furthermore, the ced-7 gene, which is a putative ABCA1 ortholog in Caenorhabditis elegans, plays a role in phagocytosis by precluding the redistribution of phagocyte receptors around the apoptotic particle (34, 35).
The expression of ABCA1 is induced by sterols (36) as well as nuclear hormone receptors, such as oxysterol receptors (LXRs) and the bile acid receptor (FXR), as heterodimers with retinoid X receptors (RXRs) (37). The promoter region contains multiple binding sites for transcription factors with roles in lipid metabolism (38–40).
Disruption of the mouse Abca1 gene results in similarly low levels of HDLs and accumulation of cholesterol in tissues (41, 42). Analyses of Abca1 –/– mice indicate that the transport of lipids from the Golgi to the plasma membrane is defective (41). However, these mice have normal secretion of cholesterol into bile, indicating that Abca1 does not play a role in this process (43). In contrast, the constitutive overexpression of Abca1 results in a protection of animals against an atherosclerotic diet (44, 45). The Wisconsin hypoalpha mutant (WHAM) chicken has been characterized as a model for Tangier disease (46) and is suspected to be mutant in Abca1 (47).
Because of the important role of ABCA1 in cholesterol transport, several groups have examined the ABCA1 gene for polymorphisms that might be associated with plasma lipid levels and cardiovascular disease. Common variation in noncoding regions of ABCA1 may significantly alter the severity of atherosclerosis, without necessarily influencing plasma lipid levels (256)
The human ABCA1 protein has been expressed in Sf9 insect cells and was found to have Mg2+-dependent ATP binding and low basal ATPase activity (257). The addition of lipid substrates did not modify the ATPase activity of ABCA1, and it was speculated that ABCA1 may be a regulatory protein or may require other protein partners for full activation.
ABCA2
The ABCA2 gene maps to chromosome 9q34.3 and is most closely related to ABCA1 (25, 48). ABCA2 is highly expressed in the brain. Given the homology to ABCA1 and its expression in the brain, it has been proposed that ABCA2 carries out similar cholesterol and phospholipid remodeling functions in neurons and glial cells.
An ovarian tumor cell line was characterized that contains an amplification of the ABCA2 gene (49). These cells are resistant to estramustine and express high levels of ABCA2 (49). Antisense treatment of these cells increases their sensitivity to the drug, supporting the idea that ABCA2 can function as a drug efflux pump.
Characterization of the full-length ABCA2 gene was performed, and antibodies to the protein demonstrate that it is localized to intracellular vesicles (258). Sterol-dependent regulation of the gene was observed, and the promoter contained several potential transcription factor-binding sites (50). The protein appears to be most highly expressed in oligodendrocytes in the brain (51).
ABCA3
The ABCA3 gene maps to chromosome 16p13.3 and is expressed as a single 7.5-kb mRNA in the lung (52, 53). Recently, it was shown that a monoclonal antibody that detects a lamellar body-specific protein in alveolar type II is directed to ABCA3 (54, 55). The lamellar bodies of type II cells produce surfactants, lipid-rich secretions that are critical to the switch of the lung from an aqueous to an air environment at birth. Surfactants also play an important role in the homeostasis of the adult lung. Surfactants are also taken up by type II cells and recycled. These data suggest that ABCA3 is directly involved in transporting lipids within the cell and participating in the production of surfactants (56).
ABCA4
The ABCA4 (ABCR) gene maps to chromosome 1p21.3 and is expressed exclusively in photoreceptors, where it believed to transport retinol (vitamin A)/phospholipid derivatives from the photoreceptor outer segment disks into the cytoplasm (52, 57, 58). These compounds are the likely substrates for ABCA4, because they stimulate the ATP hydrolysis activity of the purified protein (59). Mice lacking Abca4 show increased all-trans-retinaldehyde after light exposure, elevated phosphatidylethanolamine (PE) in outer segments, accumulation of the protonated Schiff base complex of all-trans-retinaldehyde and PE (N-retinylidene-PE), and striking deposition of a major lipofuscin fluorophore (A2-E) in retinal pigment epithelium (60). These data suggest that ABCR is an outwardly directed flippase for N-retinylidene-PE.
Mutations in the ABCA4 gene have been associated with multiple eye disorders (61). A complete loss of ABCA4 function leads to retinitis pigmentosa, whereas patients with at least one missense allele have Stargardt disease (62–64). Stargardt disease is characterized by juvenile to early adult-onset macular dystrophy with loss of central vision (65) (OMIM:248200). Nearly all patients with recessive cone rod dystrophy also have mutations in ABCA4 (64). Thus, three different recessive retinal degeneration syndromes are caused by ABCA4 mutations and are loosely correlated with the functional activity of the protein.
ABCA4 mutation carriers are also increased in frequency in age-related macular degeneration (AMD) patients (66). AMD patients display a variety of phenotypic features, including the loss of central vision, after 60 years of age. The causes of this complex trait are poorly understood, but a combination of genetic and environmental factors play a role. The abnormal accumulation of retinoids attributable to ABCA4 deficiency has been postulated to be one mechanism by which this process could be initiated. Defects in ABCA4 lead to an accumulation of retinal derivatives in the retinal pigment epithelium behind the retina. Consistent with this idea is the demonstration in ABCA4 +/– mice of light-dependent accumulation of pigmented deposits in the retinal pigment epithelium, very reminiscent of AMD (67).
ABCA5
ABCA5 is one of five ABC genes in a cluster on chromosome 17q24.3 (9, 52). A similar cluster is found on mouse chromosome 11, although the mouse cluster lacks ABCA10 and has a duplicated ABCA8 homolog (68). This cluster of ABCA genes is evolutionarily distinct from that other ABCA genes, as evidenced from phylogenetic analysis as well as analysis of intron–exon boundaries. The chromosome 17 ABCA genes have 38 exons, whereas the other ABCA genes have 50–52 exons. Therefore, it appears that all of the genes on chromosome 17 arose from an ancestral ABCA gene. This cluster is not represented in plant, nematode, or insect genomes, and there is a single ABCA5 -related gene in fish (Annilo et al, submitted). Thus ABCA5 appears to be the ancestral gene for this cluster and seems to have arisen early in vertebrate evolution.
ABCA5 is expressed as a 6.5-kb mRNA with the highest levels in pancreas, muscle, and testes (9). Neither the substrate nor the function of this gene is known.
ABCA6
ABCA6 is another member of the chromosome 17 ABCA cluster (see ABCA5 ) and is also found in the mouse genome (9, 52, 68, 69). However, the human and mouse ABCA6 genes display considerable differences, suggesting that there was a duplication of this gene and that mice and humans retained different orthologs (Annilo et al., submitted). ABCA6 is expressed as a 7.0-kb mRNA with the highest expression in the liver (9). Neither the substrate nor the function of this gene is known.
ABCA7
ABCA7 maps to chromosome 19p13.3 and is highly expressed in spleen, thymus and lymphoid cells (70, 71). ABCA7 is part of the ABCA1 / ABCA2 / ABCA3 / ABCA4 subgroup of ABCA genes. ABCA7 has 46 introns and the 3' end overlaps that of a minor histocompatabillity antigen, HA-1 (72). Intriguingly, the autoantigen SS-N, an epitope of Sjögren's syndrome, is encoded by a segment at the N terminus of the ABCA7 protein (73). Resequencing of the ABCA7 gene in 48 Japanese identified 67 single nucleotide polymorphisms, 64 of which are newly described (74). Neither the substrate nor the function of this gene is known.
ABCA8
ABCA8 is another member of the chromosome 17 ABCA cluster (see ABCA5 ) and is also found in the mouse genome (9). However, the mouse genome contains two ABCA8 -like genes that clearly arose by duplication (68) (Annilo et al., submitted). Intriguingly there are several regions of the ABCA8 and ABCA9 genes in the mouse and human genome that display evidence of gene conversion-like events. Resequencing of the ABCA8 gene from 48 Japanese people identified 88 single nucleotide polymorphisms, 78 of which are newly described (74). ABCA8 is expressed in ovary, testes, heart, and liver (Schriml and Dean, unpublished data). Neither the substrate nor the function of this gene is known.
ABCA9
ABCA9 is another member of the chromosome 17 ABCA cluster (see ABCA5 ) and is also found in the mouse genome (9, 75). ABCA9 is expressed at the highest levels in the heart and brain and induced during monocyte differentiation into macrophages and suppressed by cholesterol import (9, 75) . Neither the substrate nor the function of this gene is known.
ABCA10
ABCA10 is a member of the chromosome 17 ABCA cluster (see ABCA5 ), but the gene is absent from the mouse genome (9) (Annilo et al., submitted). ABCA10 is expressed in skeletal muscle and heart (9). Neither the substrate nor the function of this gene is known.
ABCA12
ABCA12 maps to chromosome 2q34 and is weakly expressed in the stomach (Arnould et al., in preparation). A nearly full-length sequence has also been deposited in the public databases (GenBank) by Bonner et al. The mouse gene is located on chromosome 1C3 (Dean, unpublished). Neither the substrate nor the function of this gene is known.
ABCA13
ABCA13 maps to chromosome 17p12.3 and is weakly expressed in the stomach (Annilo, in preparation). The mouse gene is located on chromosome 11A1 (Dean, unpublished). All ABCA genes are predicted to have a large extracellular loop between the first and second TM domains. However, ABCA13 contains a domain in this region that is over 3500 amino acids and is encoded by exons of 4.8 and 1.7 kb. These are among the largest exons described to date for any gene. This domain is conserved in the mouse, as are the large exons. The domain is hydrophilic and has no obvious homology to any other protein domains. The ABCA13 protein is predicted to be 5058 amino acids in length and is therefore the largest ABC protein described to date and among the largest mammalian proteins. The gene is very poorly expressed, and only 18 expressed sequence tags have been identified. Neither the substrate nor the function of this gene is known.
ABCB Genes
ABCB1
The ABCB1 (PGP/MDR1) gene maps to chromosome 7q21.1 and is the best characterized ABC drug pump. Formerly known as MDR1 or PGY1, ABCB1 was the first human ABC transporter cloned and characterized through its ability to confer a multidrug resistance phenotype to cancer cells that had developed resistance to chemotherapy drugs (76–79). ABCB1 has been demonstrated to be a promiscuous transporter of hydrophobic substrates including drugs such as colchicine, etoposide (VP16), Adriamycin, and vinblastine as well as lipids, steroids, xenobiotics, and peptides (for reviews, see Refs. 21, 80). The gene is thought to play an important role in removing toxic metabolites from cells but is also expressed in cells at the blood–brain barrier and presumably plays a role in transporting compounds into the brain that cannot be delivered by diffusion. ABCB1 also affects the pharmacology of the drugs that are substrates, and a common polymorphism in the gene affects digoxin uptake (81).
The ABCB1 protein is expressed in many secretory cell types such as kidney, liver, intestine, and adrenal gland, where the normal function is thought to involve the excretion of toxic metabolites. Mice have two closely related homologs of ABCB1 ( Abcb1a , Abcb1b ). Mice homozygous for a disrupted Abcb1a gene are phenotypically normal but are sensitive to certain neurotoxins such as ivermectin (82). Disruption of Abcb1a alone and together with Abcb1b was also accomplished (83). The double-knockout mice are viable and fertile and show similar sensitivity to ivermectin (83). These studies led to the characterization of an important role of ABCB1 in transport across the blood–brain barrier. Certain dogs of the collie breed are highly sensitive to ivermectin and have mutations in the Mdr1 gene (84).
ABCB1 is also highly expressed in hematopoietic stem cells, where it may serve to protect these cells from toxins (83, 85). ABCB1 has been shown to play a role in the migration of dendritic cells (86).
ABCB2/TAP1
The TAP1 (ABCB2) and TAP2 (ABCB3) genes are on chromosome 6p21.3 in the HLA gene complex. They are half transporters that form a heterodimer that serves to transport peptides into the ER, where they can be complexed with class I HLA molecules for presentation on the cell surface (87–89). TAP expression is required for the stable expression of class I proteins (90). In vitro systems have been used to define the substrate specificity of the transporter (91, 92). These studies have shown that the TAP complex preferentially transports 9–12 amino acid peptides (93) with a preference for Phe, Leu, Arg, and Tyr at the C terminus, similar to the specificity of the HLA class I proteins (93, 94). Tap1-deficient mice are deficient in antigen presentation and surface class I molecules and lack CD8+ cells (95).
Several DNA viruses such as herpes simplex virus express molecules that interfere with antigen expression by disrupting the function of the TAP complex (96–99). In addition, tumor cell lines have been described that are mutated and deficient in TAP activity (100). Patients with inherited immunodeficiency because of TAP1 mutations have been described (101).
ABCB3/TAP2
The TAP2 (ABCB3) gene maps to chromosome 6p21.3 and functions as a heterodimer with TAP1 (see TAP1). A family with recessive inheritance of class I HLA deficiency was described that has a nonsense mutation in the TAP2 gene (102).
ABCB4
The ABCB4 (MDR3/PGY3) gene maps to 7q21.1, adjacent to the ABCB1 (PGP/MDR1) gene, and encodes a full transporter with high homology to ABCB1 . These genes clearly arose by duplication, although the function of ABCB4 is very different from ABCB1 . ABCB4 is principally expressed in the bile cannilicular membrane of the liver, but is also found in the heart, muscle and in B cells. Disruption of the gene in the mouse resulted in liver pathology because of a deficiency in fatty acid secretion in bile (103). In vitro experiments confirm that ABCB4 can transport phophatidylcholine from the inner to the outer leaflet of the membrane (104, 105). Mutations in this gene cause PFIC3 (106, 107) and are associated with intrahepatic cholestasis of pregnancy (108, 109).
ABCB5
The ABCB5 gene maps to 7p21.1 and encodes a full transporter molecule (52) (Allikmets, unpublished). The gene is expressed as a 7.5-kb transcript in all cells and has no described function.
ABCB6
The ABCB6 gene maps to chromosome 2q35, and the protein is localized to the mitochondria (52, 110). It is closely related to the ABCB7 protein: both are half transporters. The ABCB6 gene, similar to ABCB7 , can complement yeast cells that are defective in the ATM1 gene, a mitochondrial ABC gene that is involved in the transport of a precursor of the Fe/S cluster from mitochondria to the cytosol (110).
ABCB7
The ABCB7 gene maps to chromosome Xq21–q22, and the protein is localized to the mitochondria (52, 111). It is closely related to the ABCB6 gene, both of which are half transporters. The human ABCB7 gene can complement yeast cells that are defective in the ATM1 gene, a mitochondrial ABC gene that is involved in the transport and/or maturation of a precursor of the Fe/S cluster from mitochondria to the cytosol (111, 112). The ABCB7 gene is mutated in patients with X-linked sideroblastic anemia and ataxia (XLSA/A) (112, 113). XLSA/A is a recessive disorder characterized by infantile to early childhood onset of non-progressive cerebellar ataxia and mild anemia with hypochromia and microcytosis.
An I400M variant in ABCB7 was identified in a predicted TM segment of the ABCB7 gene in patients from an XLSA/A family. The mutation was shown to segregate with the disease in the family and was not detected in at least 600 chromosomes of general population controls. Introduction of the corresponding mutation into the S. cerevisiae ATM1 gene resulted in a partial loss of function of the yeast Atm1 protein (112). A second family with an E433K mutation was also identified. The analogous E433K mutation in the yeast ATM1 gene (D398K) also results in loss of function, as assessed by cytosolic Fe/S protein maturation (113).
ABCB8
The ABCB8 (M-ABC1) gene maps to 7q36.1 and encodes a half transporter protein located in the mitochondria, although its function is unknown (52, 114).
ABCB9
The ABCB9 gene maps to 12q24.31 and encodes a half transporter protein located in the lysosomes, the function of which is unknown (52, 115).
ABCB10
The ABCB10 (M-ABC2) gene maps to 1q42.13 and encodes a half transporter protein located in the mitochondria, although its function is unknown (48, 116).
ABCB11
The ABCB11 (BSEP/SPGP) gene maps to 2q24.3 and encodes a full transporter protein located principally, if not exclusively, in the liver (117, 118). The protein localizes to bile canalicular membrane of the liver and participates in the secretion of bile salts such as taurocholate (118). ABCB11 protein is localized to vesicles within liver cells lining the bile duct.
Mutations in ABCB11 are found in patients with progressive familial intrahepatic cholestasis, type 2 (PFIC2) (119). Disruption of the murine Abcb11 gene results in intrahepatic cholestasis. However, the phenotype is less severe and indicates that mice display compensatory changes (120). Analysis of the ABCB11 promoter showed a farnesoid X receptor (FXR)-responsive element (FXRE) at position –180 (121). The FXR functions as a heterodimer with the retinoid X receptorα (RXRα) and can be activated by the bile salt chenodeoxycholic acid. Thus, similar to several ABC genes, ABCB11 is regulated by its ligand.
ABCC Genes
ABCC1
The ABCC1 (MRP1) gene maps to chromosome 16p13.1 and is expressed in tumor cells (122). ABCC1 is adjacent to the ABCC6 gene, and one of these genes undoubtedly arose by gene duplication. It encodes a full transporter that is the principal transporter of glutathione-linked compounds from cells. The ABCC1 gene was identified in the small cell lung carcinoma cell line NCI-H69, a multidrug-resistant cell that does not overexpress ABCB1 (123). The ABCC1 pump confers resistance to doxorubicin, daunorubicin, vincristine, colchicines, and several other compounds, very similar profile to that of ABCB1 (124). However, unlike ABCB1, ABCC1 transports drugs that are conjugated to glutathione by the glutathione reductase pathway (12, 122, 125–127).
Disruption of the Abcc1 gene demonstrated that it is not essential for viability or fertility (128, 129). However, these mice do display an impaired inflammatory response and they are hypersensitive to the anticancer drug etoposide.
ABCC1 can transport leukotrienes such as leukotriene C4 (LTC4). LTC4 is an important signaling molecule for the migration of dendritic cells. Migration of dendritic cells from the epidermis to lymphatic vessels is defective in Abcc1 –/– mice, implicating a role for LTC4 in the response of dendritic cells to chemokines (130). The ABCC1 protein is thought to play both a role in protecting cells from chemical toxicity and oxidative stress and to mediate inflammatory responses involving cysteinyl leukotrienes (122).
ABCC2
The ABCC2 (MRP2/cMOAT) gene maps to chromosome 10q24 and is expressed in canalicular cells in the liver (52, 131). It functions as the major exporter of organic anions from the liver into the bile. The role of ABCC2 in organic ion transport was first elucidated by the discovery that this gene is mutated in the TR-rat, a rat strain that displays jaundice and a deficiency in organic ion transport (132). Subsequently, it was found that the gene is also mutated in patients with Dubin–Johnson syndrome, a human disorder of organic ion transport (133, 134). ABCC2 overexpression can confer drug resistance to cells, but the physiological importance of this observation is not clear (12, 124, 135).
The localization of the ABCC2 protein on the membrane of the bile canaliculus is dependent of the expression of the radixin ( RDX ) product. Rdx –/– mice have increased bilirubin and develop liver injury, similar to Dubin–Johnson patients (136).
ABCC3
The ABCC3 (MRP3) gene maps to 17q21.3 and is expressed primarily in the liver (52, 131). Similar to ABCC2, ABCC3 can confer the ability to efflux organic ions, and cells become resistant to certain cytotoxic compounds (137, 138).
ABCC4
The ABCC4 (MRP4, MOATB) gene maps to 13q32 and is expressed at low levels in many cell types and tissues (52, 131, 139). Overexpression and amplification of the ABCC4 gene is found in cell lines resistant to nucleoside analogues such as azidothymidine monophosphate (140). Transfection of ABCC4 into cells confers resistance to these compounds (140). Thus, ABCC4 may be an important factor in the resistance to nucleoside analogues. Because these drugs are important antiviral and anticancer agents, this has importance in therapies for human immunodeficiency virus 1 infection and other disease.
ABCC5
The ABCC5 (MRP5/MOATC) gene maps to 3q27 and is ubiquitously expressed in tissues and cells (52, 131, 141). It is closely related to the ABCC4 gene and also confers resistance to nucleoside analogues (142).
ABCC6
The ABCC6 (MRP6) gene maps to 16p13.1, adjacent to the ABCC1 gene. The gene is principally expressed in the liver and kidney. ABCC6 is mutated in pseudoxanthoma elasticum, a recessive genetic disorder characterized by calcification of the connective fibers of the skin, ocular bleeding, and cardiovascular disease (143–147). Several pseudogenes of ABCC6 have been identified that also map to 16p (148, 149).
Expression of the human ABCC6 protein in Sf9 insect cells demonstrated that the protein is present in isolated membranes and can transport glutathione conjugates including LTC4 (150). Organic anions inhibit transport, and the expression of three missense mutations found in PXE patients abolished transport activity. Expression and purification of the rat Abcc6 protein demonstrated Mg2+-dependent trapping of 8-azido-ATP. However, stimulation of nucleotide binding could not be demonstrated by glutathione conjugates (151), and glutathione conjugate transport by the purified rat gene was not detected (152).
Analysis of the ABCC6 gene for variants has identified a number of common polymorphisms including missense alleles (148). One of these variants, R1268Q, is associated with plasma triglyceride and HDL levels (148). The R1141X mutation is the most prevalent ABCC6 mutation in PXE patients of European descent, and this variant has been found at levels approaching 1% in these populations. An association of this variant with premature atherosclerotic vascular disease has been reported (153).
ABCC7/CFTR
The CFTR (ABCC7) gene maps to chromosome 7q31.2 and is a protein kinase A-dependent chloride channel expressed in exocrine tissues such as the sweat duct, pancreas, intestine, and kidney. The gene is mutated in the recessive genetic disease cystic fibrosis (154–156).
Cystic fibrosis (CF) is the most common fatal childhood disease in Caucasian populations and is characterized by defective exocrine activity of the lung, pancreas, sweat ducts, and intestine (11, 157). The disease is found at frequencies ranging from 1/900 to 1/2500. This corresponds to a carrier frequency of 1/15 to 1/25. The disease is much less common in African and Asian populations, where carrier frequencies of 1/100 to 1/200 have been estimated. In most populations, the disease frequency correlates with the frequency of the major allele of the CFTR gene, a deletion of 3 base pairs (ΔF508) (158). However, at least two other populations have high-frequency CFTR alleles. The W1282X allele is found on 51% of the alleles in the Ashkenazi Jewish population, and the 1677delTA allele has been found at a high frequency in Georgians and is also present at an elevated level in Turkish and Bulgarian populations. This has led several groups to hypothesize that these alleles arose through selection of an advantageous phenotype in the heterozygotes (159). It is through CFTR that some bacterial toxins, such as cholera toxin, and those from Escherichia coli cause increased fluid flow in the intestine and result in diarrhea. Therefore, several researchers have proposed that the CF mutations have been selected for in response to this disease(s). This hypothesis is supported by studies showing that: (a) CF homozygotes indeed fail to secrete chloride ions in response to a variety of stimulants; and (b) mice in heterozygous null animals showed reduced intestinal fluid secretion in response to cholera toxin (160). CFTR is also the receptor for Salmonella typhi toxin and has an implied functional role in the innate immunity to Pseudomonas aeruginosa (161).
Cftr –/– mice display many of the hallmarks of the human disease, including defects in the bowel and male reproductive tract (162, 163). A mouse model of the ΔF508 mutation has also bee generated (164).
Patients with two severe CFTR alleles such as ΔF508 typically display severe disease with inadequate secretion of pancreatic enzymes leading to nutritional deficiencies, bacterial infections of the lung, and obstruction of the vas deferens leading to male infertility (165, 166). Patients with at least one partially functional allele display enough residual pancreatic function to avoid the major nutritional and intestinal deficiencies (167), and subjects with very mild alleles display only congenital absence of the vas deferens with none of the other symptoms of CF (168, 169). Recently, heterozygotes of CF mutations have been found to have an increased frequency of pancreatitis (170) and bronchiectasis (171). Thus, there is a spectrum of severity in the phenotypes caused by this gene that is inversely related with the level of CFTR activity. Clearly, other modifying genes and the environment also affect disease severity, particularly the pulmonary phenotypes.
Several research groups have approached gene therapy in the lung as a potential treatment for CF. This approach has proved extremely difficult and may require more detailed insight into the cell types that express CFTR in the lung (reviewed in Refs. 172, 173).
The identification of the CFTR gene led to expression of both the wild-type and mutant forms of the protein and to considerable insight into its function, regulation, and ability to regulate other ion channels (reviewed in Refs 174–176). Although a large number of CF mutations occur in the NBFs and function to inactivate the protein, a number of CFTR alleles also cause misprocessing of the protein (reviewed in Ref. 177). The CFTR protein is unusual amongst ABC genes in having a large, hydrophilic domain after the first NBF (154). This domain, the R domain, is phosphorylated by cAMP-dependent kinases and serves to regulate the activity of the channel (reviewed in Ref. 178).
ABCC8
The ABCC8 (SUR1) gene maps to chromosome 11p15.1 and encodes a full transporter molecule. The gene is closely related to ABCC9 (SUR2). The ABCC8 gene codes for a high-affinity receptor for the drug sulfonylurea. Sulfonylureas are a class of drugs widely used to increase insulin secretion in patients with non-insulin-dependent diabetes. These drugs bind to the ABCC8 protein and inhibit an associated potassium channel K(ATP). Familial persistent hyperinsulinemic hypoglycemia of infancy is an autosomal recessive disorder in which subjects display unregulated insulin secretion. The disease was mapped to 11p15–p14 by linkage analysis, and mutations in the ABCC8 gene are found in PHHI families (179). Sur1 –/– mice also lack K(ATP) channels; however, they show normal glucose levels, suggesting that compensatory pathways are present in mice (180).
The ABCC8 gene has also been implicated in insulin response in Mexican-American subjects (181) and in type II diabetes in French Canadians (182) but not in a Scandinavian cohort (183).
ABCC9
The ABCC9 (SUR2) gene maps to 12p12.1 and is closely related to the ABCC8 (SUR1) gene on chromosome 11. ABCC9 shows low-affinity binding to sulfonylurea and is the primary regulator of K(ATP) channels in muscle. Sur2 –/– mice display enhanced insulin-stimulated glucose uptake in skeletal muscle (184).
ABCC10
The ABCC10 gene (MRP7) maps to 6p21.1 and groups with the other ABCC1 -related genes ( ABCC2 , ABCC3 , ABCC4 , ABCC5 , ABCC6 , ABCC11 , and ABCC12 ) (52). However, the function of ABCC10 is not known.
ABCC11
The ABCC11 (MRP8) gene maps to 16q12.1 in a cluster with the ABCC12 (MRP9) gene (185–187). A human T cell leukemia cell line that is resistant to nucleoside drugs overexpresses ABCC11 (188). The mouse appears to have only a single gene in this cluster, indicating that the duplication occurred relatively recently (Dean, unpublished).
ABCD Genes
ABCD1
The ABCD1 (ALD) gene maps to Xq28 and expresses a peroxisomally located half transporter that is mutated in adrenoleukodystrophy (ALD). X-ALD is an X-linked recessive disorder characterized by neurodegenerative phenotypes with onset typically in late childhood (189). Adrenal deficiency commonly occurs, and the presentation of ALD is highly variable. Childhood ALD, adrenomyeloneuropathy, and adult onset forms are recognized, but there is no apparent correlation to ABCD1 alleles (190). Female heterozygotes can display symptoms including spastic paraparesis and peripheral neuropathy (191).
More than 406 mutations have been documented in the ABCD1 gene and a database of ALD mutations has been created (190) (http://www.x-ald.nl). Although most mutations are point mutations, several large intragenic deletions have also been described (192). A contiguous gene syndrome, contiguous ABCD1 DXS1357E deletion syndrome (CADDS), has been described that includes ABCD1 and the adjacent DXS1357E gene. These patients present with symptoms at birth, as opposed to X-ALD, which present after 3 years of age (193).
ALD patients have an accumulation of unbranched saturated fatty acids, with a chain length of 24–30 carbons, in the cholesterol esters of the brain and in adrenal cortex. The ALD protein is located in the peroxisome, where it is believed to be involved in the transport of very long chain fatty acids (VLCFAs). A treatment consisting of erucic acid, a C22 monounsaturated fat, and oleic acid, a C18 monounsaturated fat (Lorenzo's oil), was developed that results in a normalization of the VLCFA levels in the blood of patients but does not appear to dramatically slow the progression of the disease (194). This is probably because the treatment fails to lower fatty acid levels in the brain (195). An Abcd1 –/– mouse has been generated, and the animals display accumulation of VLCFAs in kidney and brain; however, they do not show the severe neurological abnormalities of the childhood cerebral form of X-ALD (196, 197). The mice do show evidence of a late-onset neurological disorder characterized by slower nerve conduction and myelin and axonal anomalies detectable in the spinal cord and sciatic nerve (198).
ABCD1 is one of four related peroxisomal transporters that are found in the human genome, the others being ABCD2, ABCD3, and ABCD4. These genes are highly conserved in evolution, and a pair of homologous genes is present in the yeast genome, PXA1 and PXA2 . The PXA2 gene has been demonstrated to transport long-chain fatty acids (199, 200). A defective pxa1 gene in Arabidopsis thaliana results in defective import of fatty acids into the peroxisome (201).
ABCD2
The ABCD2 (ALDR) gene maps to chromosome 12q11 and encodes a 741-amino acid half transporter that is 66% identical at the amino acid level with ABCD2 (202, 203). The ABCD2 protein is expressed in peroxisomes and is particularly abundant in the brain and adrenal gland (202). The ABCD1 and ABCD2 genes share the same exon/intron structure, further evidence that they are closely related (204). Overexpression of the ABCD2 gene in cells from X-ALD patients at least partially restores the impaired peroxisomal β-oxidation in fibroblasts (205). The ABCD2 gene is induced by fibrates (cholesterol-lowering drugs) in a peroxisome proliferator-activated receptor (PPARα)-dependant fashion, providing a potential therapeutic strategy to treat X-ALD (206).
ABCD3
The ABCD3 (PMP70/PXMP1) gene maps to chromosome 1p21–p22 and encodes a peroxisomal protein. Although mutations in ABCD3 were found in two patients with Zellweger syndrome (207), further evidence does not support a role for ABCD3 in this disorder (208).
ABCD4
The ABCD4 (PXMP1L/P70R/PMP69) gene maps to chromosome 14q24.3 and encodes the fourth peroxisomal half transporter (52, 209, 210). ABCD4 shares 25–27% amino acid identity with the ABCD1, ABCD2, and ABCD3 proteins. The gene contains 19 exons and spans approximately 16 kb and encodes several differentially spliced mRNAs (211).
ABCE Genes
ABCE1
The ABCE1 (RNS4I) gene maps to 4q31 (52, 212) and encodes a protein with two ATP-binding domains with high homology to other ABC genes but no TM domains. Along with the genes in the ABCF subfamily, ABCE genes are cytosolically expressed ABC genes that are not membrane transporters. However, they all clearly possess ABC-type NBFs and are therefore included in the gene superfamily (3, 5).
ABCE1 inhibits the RNaseL protein, a ribonuclease that is activated by interferons (213). The ABCE1 gene is expressed as 2.4- and 3.8-kb mRNAs in all tissues (214). ABCE1 has been found recently to be essential for the assembly of immature human immunodeficiency virus capsids (215).
ABCF Genes
ABCF1
The ABCF1 (ABC50) gene is localized to chromosome 6p21.33 inside the class I HLA complex and encodes a protein with two ATP-binding domains and no TM domains (52). The gene is activated by tumor necrosis factor-α stimulation of cells (216). The human genome contains three ABCF genes of unknown function. The yeast ABCF homologs include the GCN20 gene, which codes for a protein required for the activation of a kinase that phosphorylates the translation initiation factor eIF2 (15). The ABCF1 protein associates with human ribosomes and copurifies with eIF2, suggesting that it performs an analogous function in human cells (16).
ABCG Genes
ABCG1
The ABCG1 gene is located on chromsome 21q22.3 and encodes a half transporter (17, 217, 218). Similar to all ABCG family genes, the NBF is at the N terminus, and the TM domains are at the C terminus, the opposite orientation of all other eukaryotic ABC genes. The ABCG1 gene is 31% identical to the Drosophila white gene, a transporter of eye pigment precursors. It is most closely related to the ABCG4 gene, and these two genes are the only human ABCG genes that share a conserved intron location, indicating that they arose from a recent duplication (219).
The ABCG1 gene is induced by cholesterol in monocyte-derived macrophages during cholesterol influx mediated by acetylated low-density lipoprotein (18). This suggests that, similar to ABCG5 and ABCG8 , ABCG1 is involved in cholesterol efflux (33, 220). ABCG1 contains a TATA-less, GC-rich promoter that contains silencing elements that can mediate transcriptional repression (221). Multiple alternative transcripts affecting the N terminus of the protein have been identified, as has a second promoter region. Both promoters were found to be responsive to hydroxycholesterol and retinoic acid in macrophages.
The mouse Abcg1 gene maps to chromosome 17A2-B and has 97% identity to the human locus (217, 218).
ABCG2
The ABCG2 (MXR/BCRP/ABCP) gene maps to chromosome 4q22 and encodes a half transporter with a NBF–TM orientation (52, 222). Analysis of cell lines resistant to mitoxantrone that do not overexpress ABCB1 or ABCC1 led several laboratories to identify the ABCG2 gene as a drug transporter (222-224). ABCG2 confers resistance to anthracycline anticancer drugs and is amplified or involved in chromosomal translocations in cell lines selected with topotecan, mitoxantrone, or doxorubicin treatment. It is suspected that ABCG2 functions as a homodimer because transfection of the gene into cells confers resistance to chemotherapeutic drugs (225). Variations at residue 482 of ABCG2 are found in many resistant cell lines, and the alteration of the wild-type arginine at this position for either threonine or glycine imparts the ability to transport rhodamine and alters the substrate specificity (226).
ABCG2 can also transport several dyes, such as rhodamine and Hoechst 33462, and the gene is highly expressed in a subpopulation of hematopoetic stem cells (side population) that stain poorly for these dyes (227–229). However, the normal function of the gene in these cells is unknown. ABCG2 is highly expressed in the trophoblast cells of the placenta (230). This suggests that the pump is responsible either for transporting compounds into the fetal blood supply or removing toxic metabolites (231). The gene is also expressed in the intestine, and inhibitors could be useful in making substrates orally available.
In mouse cells from animals deficient in the Abcb1 , Abcb1a , and Abcc1 genes, exposure to mitoxantrone, topotecan, or doxorubicin results in amplification of the Abcg2 gene (20). This strongly suggests that ABCG2 is one of three major transporter genes involved in drug resistance in mammalian cells. Inhibitors of ABC drug transporters represent a potential strategy for preventing the development of drug-resistant tumors (21). Effective inhibitors of ABCG2, such as fumitrimorgin C, a natural product from Aspergillus, and GF120918 have been described (223, 232–234).
ABCG3
The Abcg3 (Abcp2) gene maps to chromosome 4 and is highly related to ABCG2 . The gene is principally expressed in murine hematopoeitic cells and has no ortholog in the human genome, although other rodents appear to have a orthologous sequence (22). The gene has an unusual NBF domain that has alternative residues in several conserved positions, suggesting that it might either fail to bind or hydrolyze ATP (22).
ABCG4
The ABCG4 gene maps to 11q23 and expresses a half transporter protein that is highly related to ABCG1 . ABCG4 has the same intron/exon structure as ABCG1 , suggesting that they arose by a relatively recent gene duplication event (219). The gene is primarily expressed in the brain; however, there are several alternative transcripts that are specifically expressed in either hematopoetic cells and in the lung (219). Similar to ABCG1 , ABCG4 is also induced by oxysterols and retinoids (235). The murine Abcg4 gene is 98% identical to the human gene and is highly expressed in the brain, spleen, eye, and bone marrow (236).
ABCG5
The ABCG5 gene maps to chromosome 2p21 and is adjacent to and arranged head-to-head with the ABCG8 gene (237–239) (Figure 6). Both of these genes are mutated in families with sitosterolemia, a disorder characterized by defective transport of plant and fish sterols and cholesterol (238–243). Most likely, the two half transporters form a functional heterodimer, and they appear to be regulated by the same promoter (244).
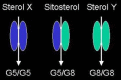
Patients mutated in either ABCG5 or ABCG8 have similarly elevated levels of sitosterol, suggesting that it is the heterodimer that is the principal transporter of sitosterol (245). However, Asian sitosterolemia patients have almost exclusive mutations in ABCG5 , and Caucasian patients have mutations in ABCG8 (245–247). This suggests that there are independent functions of the two genes, and that they may also form heterodimers to transport some of the wide variety of non-cholesterol sterols found in plants and shellfish.
ABCG8
The ABCG8 gene is adjacent to the ABCG5 gene on chromosome 2p21, and the two genes are coordinately induced by cholesterol (238) (Figure 6). The levels of sterols in blood were demonstrated to be highly heritable, and at least two variants in ABCG8 (D19H and T400K) were shown to be associated with lower concentrations of sterols in parents and their offspring (248). Several additional frequent missense variants in the ABCG8 gene were also described (245, 249), suggesting that both ABCG5 and ABCG8 are functionally polymorphic, perhaps in response to selection based on dietary sterol exposure.
References
- 1.
- Higgins CF . ABC transporters: from micro-organisms to man. Annu Rev Cell Biol. 1992;8:67–113. [PubMed: 1282354]
- 2.
- Childs S , Ling V . The MDR superfamily of genes and its biological implications. Important Adv Oncol. 1994:21–36. [PubMed: 7911445]
- 3.
- Dean M , Allikmets R . Evolution of ATP-binding cassette transporter genes. Curr. Opin. Genet. Dev. 1995;5:779–785. [PubMed: 8745077]
- 4.
- Hyde S C , Emsley P , Hartshorn M J , Mimmack M M , Gileadi U , Pearce S R , Gallagher M P , Gill D R , Hubbard R E , Higgins C F . Structural model of ATP-binding proteins associated with cystic fibrosis, multidrug resistance and bacterial transport. Nature. 1990;346:362–5. [PubMed: 1973824]
- 5.
- Dean M , Rzhetsky A , Allikmets R . The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001;11:1156–66. [PubMed: 11435397]
- 6.
- Venter J C , Adams M D , Myers E W , Li P W , Mural R J , Sutton G G , Smith H O , Yandell M , Evans C A , Holt R A , Gocayne J D , Amanatides P , Ballew R M , Huson D H , Wortman J R , Zhang Q , Kodira C D , Zheng X H , Chen L , Skupski M , Subramanian G , Thomas P D , Zhang J , Gabor Miklos G L , Nelson C , Broder S , Clark A G , Nadeau J , McKusick V A , Zinder N , Levine A J , Roberts R J , Simon M , Slayman C , Hunkapiller M , Bolanos R , Delcher A , Dew I , Fasulo D , Flanigan M , Florea L , Halpern A , Hannenhalli S , Kravitz S , Levy S , Mobarry C , Reinert K , Remington K , Abu-Threideh J , Beasley E , Biddick K , Bonazzi V , Brandon R , Cargill M , Chandramouliswaran I , Charlab R , Chaturvedi K , Deng Z , Francesco V D , Dunn P , Eilbeck K , Evangelista C , Gabrielian A E , Gan W , Ge W , Gong F , Gu Z , Guan P , Heiman T J , Higgins M E , Ji R R , Ke Z , Ketchum K A , Lai Z , Lei Y , Li Z , Li J , Liang Y , Lin X , Lu F , Merkulov G V , Milshina N , Moore H M , Naik A K , Narayan V A , Neelam B , Nusskern D , Rusch D B , Salzberg S , Shao W , Shue B , Sun J , Wang Z Y , Wang A , Wang X , Wang J , Wei M H , Wides R , Xiao C , Yan C . et al. The sequence of the human genome. Science. 2001;291:1304–1351. [PubMed: 11181995]
- 7.
- Lander E S , Linton L M , Birren B , Nusbaum C , Zody M C , Baldwin J , Devon K , Dewar K , Doyle M , FitzHugh W , Funke R , Gage D , Harris K , Heaford A , Howland J , Kann L , Lehoczky J , LeVine R , McEwan P , McKernan K , Meldrim J , Mesirov J P , Miranda C , Morris W , Naylor J , Raymond C , Rosetti M , Santos R , Sheridan A , Sougnez C , Stange-Thomann N , Stojanovic N , Subramanian A , Wyman D , Rogers J , Sulston J , Ainscough R , Beck S , Bentley D , Burton J , Clee C , Carter N , Coulson A , Deadman R , Deloukas P , Dunham A , Dunham I , Durbin R , French L , Grafham D , Gregory S , Hubbard T , Humphray S , Hunt A , Jones M , Lloyd C , McMurray A , Matthews L , Mercer S , Milne S , Mullikin J C , Mungall A , Plumb R , Ross M , Shownkeen R , Sims S , Waterston R H , Wilson R K , Hillier L W , McPherson J D , Marra M A , Mardis E R , Fulton L A , Chinwalla A T , Pepin K H , Gish W R , Chissoe S L , Wendl M C , Delehaunty K D , Miner T L , Delehaunty A , Kramer J B , Cook L L , Fulton R S , Johnson D L , Minx P J , Clifton S W , Hawkins T , Branscomb E , Predki P , Richardson P , Wenning S , Slezak T , Doggett N , Cheng J F , Olsen A , Lucas S , Elkin C , Uberbacher E , Frazier M . et al. Initial sequencing and analysis of the human genome. International Human Genome Sequencing Consortium. Nature. 2001;409:860–921. [PubMed: 11237011]
- 8.
- Broccardo C , Luciani M , Chimini G . The ABCA subclass of mammalian transporters. Biochim Biophys Acta. 1999;1461:395–404. [PubMed: 10581369]
- 9.
- Arnould I , Schriml L , Prades C , Lachtermacher-Triunfol M , Schneider T , Maintoux C , Lemoine C , Debono D , Devaud C , Naudin L , Bauché S , Annat M , Allikmets R , Patrice Denèfle P , Rosier M , Dean M . Identification and characterization of a cluster of five new ATP-binding cassette transporter genes on human chromosome 17q24: a new sub-group within the ABCA sub-family. GeneScreen. 2001;1:157–164.
- 10.
- Anjard C , Consortium D S , Loomis W F . Evolutionary analyses of ABC transporters of Dictyostelium discoideum Eukaryotic Cell (in press); 2002. [PMC free article: PMC117992] [PubMed: 12456012]
- 11.
- Quinton P M . Physiological basis of cystic fibrosis: a historical perspective. Physiol Rev. 1999;79:S3–S22. [PubMed: 9922374]
- 12.
- Borst P , Evers R , Kool M , Wijnholds J . A family of drug transporters: the multidrug resistance-associated proteins. J Natl Cancer Inst. 2000;92:1295–302. [PubMed: 10944550]
- 13.
- Bakos E , Evers R , Calenda G , Tusnady G E , Szakacs G , Varadi A , Sarkadi B . Characterization of the amino-terminal regions in the human multidrug resistance protein (MRP1). J Cell Sci. 2000;113:4451–4461. [PubMed: 11082039]
- 14.
- Shani N , Valle D . Peroxisomal ABC transporters. Methods Enzymol. 1998;292:753–76. [PubMed: 9711597]
- 15.
- Marton M J , Vazquez de Aldana C R , Qiu H , Chakraburtty K , Hinnebusch A G . Evidence that GCN1 and GCN20, translational regulators of GCN4, function on elongating ribosomes in activation of eIF2α kinase GCN2. Mol Cell Biol. 1997;17:4474–89. [PMC free article: PMC232301] [PubMed: 9234705]
- 16.
- Tyzack J K , Wang X , Belsham G J , Proud C G . ABC50 interacts with eukaryotic initiation factor 2 and associates with the ribosome in an ATP-dependent manner. J Biol Chem. 2000;275:34131–9. [PubMed: 10931828]
- 17.
- Chen H , Rossier C , Lalioti M D , Lynn A , Chakravarti A , Perrin G , Antonarakis S E . Cloning of the cDNA for a human homologue of the Drosophila white gene and mapping to chromosome 21q22. 3, Am J Hum Genet. 1996;59:66–75. [PMC free article: PMC1915121] [PubMed: 8659545]
- 18.
- Klucken J , Buchler C , Orso E , Kaminski W E , Porsch-Ozcurumez M , Liebisch G , Kapinsky M , Diederich W , Drobnik W , Dean M , Allikmets R , Schmitz G . ABCG1 (ABC8), the human homolog of the Drosophila white gene, is a regulator of macrophage cholesterol and phospholipid transport. Proc Natl Acad Sci U S A. 2000;97:817–822. [PMC free article: PMC15414] [PubMed: 10639163]
- 19.
- Klein I , Sarkadi B , Varadi A . An inventory of the human ABC proteins. Biochim Biophys Acta. 1999;1461:237–62. [PubMed: 10581359]
- 20.
- Allen J D , Brinkhuis R F , Wijnholds J , Schinkel A H . The mouse Bcrp1/Mxr/Abcp gene: amplification and overexpression in cell lines selected for resistance to topotecan, mitoxantrone, or doxorubicin. Cancer Res. 1999;59:4237–41. [PubMed: 10485464]
- 21.
- Gottesman M M , Fojo T , Bates S E . Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer. 2002;2:48–58. [PubMed: 11902585]
- 22.
- Mickley L , Jain P , Miyake K , Schriml L M , Rao K , Fojo T , Bates S , Dean M . An ATP-binding cassette gene (ABCG3) closely related to the multidrug transporter ABCG2 (MXR/ABCP) has an unusual ATP-binding domain. Mamm Genome. 2001;12:86–88. [PubMed: 11178751]
- 23.
- Myers E W , Sutton G G , Delcher A L , Dew I M , Fasulo D P , Flanigan M J , Kravitz S A , Mobarry C M , Reinert K H , Remington K A , Anson E L , Bolanos R A , Chou H H , Jordan C M , Halpern A L , Lonardi S , Beasley E M , Brandon R C , Chen L , Dunn P J , Lai Z , Liang Y , Nusskern D R , Zhan M , Zhang Q , Zheng X , Rubin G M , Adams M D , Venter J C . A whole-genome assembly of Drosophila. Science. 2000;287:2196–204. [PubMed: 10731133]
- 24.
- Wu C T , Budding M , Griffin M S , Croop J M . Isolation and characterization of Drosophila multidrug resistance gene homologs. Mol Cell Biol. 1991;11:3940–8. [PMC free article: PMC361188] [PubMed: 2072901]
- 25.
- Luciani M F , Denizot F , Savary S , Mattei M G , Chimini G . Cloning of two novel ABC transporters mapping on human chromosome 9. Genomics. 1994;21:150–9. [PubMed: 8088782]
- 26.
- Bodzioch M , Orso E , Klucken J , Langmann T , Bottcher A , Diederich W , Drobnik W , Barlage S , Buchler C , Porsch-Ozcurumez M , Kaminski W E , Hahmann H W , Oette K , Rothe G , Aslanidis C , Lackner K J , Schmitz G . The gene encoding ATP-binding cassette transporter 1 is mutated in Tangier disease. Nat Genet. 1999;22:347–51. [PubMed: 10431237]
- 27.
- Brooks-Wilson A , Marcil M , Clee S M , Zhang L H , Roomp K , van Dam M , Yu L , Brewer C , Collins J A , Molhuizen H O , Loubser O , Ouelette B F , Fichter K , Ashbourne-Excoffon K J , Sensen C W , Scherer S , Mott S , Denis M , Martindale D , Frohlich J , Morgan K , Koop B , Pimstone S , Kastelein J J , Hayden M R . et al. Mutations in ABC1 in Tangier disease and familial high-density lipoprotein deficiency. Nat Genet. 1999;22:336–45. [PubMed: 10431236]
- 28.
- Rust S , Rosier M , Funke H , Real J , Amoura Z , Piette J C , Deleuze J F , Brewer H B , Duverger N , Denefle P , Assmann G . Tangier disease is caused by mutations in the gene encoding ATP-binding cassette transporter 1. Nat Genet. 1999;22:352–5. [PubMed: 10431238]
- 29.
- Remaley A T , Rust S , Rosier M , Knapper C , Naudin L , Broccardo C , Peterson K M , Koch C , Arnould I , Prades C , Duverger N , Funke H , Assman G , Dinger M , Dean M , Chimini G , Santamarina-Fojo S , Fredrickson D S , Denefle P , Brewer H B Jr. Human ATP-binding cassette transporter 1 (ABC1): genomic organization and identification of the genetic defect in the original Tangier disease kindred. Proc Natl Acad Sci U S A. 1999;96:12685–90. [PMC free article: PMC23050] [PubMed: 10535983]
- 30.
- Young S G , Fielding C J . The ABCs of cholesterol efflux. Nat Genet. 1999;22:316–8. [PubMed: 10431227]
- 31.
- Marcil M , Brooks-Wilson A , Clee S M , Roomp K , Zhang L H , Yu L , Collins J A , van Dam M , Molhuizen H O , Loubster O , Ouellette B F , Sensen C W , Fichter K , Mott S , Denis M , Boucher B , Pimstone S , Genest J Jr., Kastelein J J , Hayden M R . Mutations in the ABC1 gene in familial HDL deficiency with defective cholesterol efflux. Lancet. 1999;354:1341–6. [PubMed: 10533863]
- 32.
- Ishii J , Nagano M , Kujiraoka T , Shihara M , Egashira T , Takada D , Tsuji M , Hattori H , Emi M . Clinical variant of Tangier disease in Japan: mutation of the ABCA1 gene in hypoalphalipoproteinemia with corneal lipidosis. J Hum Genet. 2002;47:366–9. [PubMed: 12111371]
- 33.
- Dean M , Hamon Y , Chimini G . The human ATP-binding cassette (ABC) transporter superfamily. J Lipid Res. 2001;42:1007–17. [PubMed: 11441126]
- 34.
- Wu Y C , Horvitz H R . The C. elegans cell corpse engulfment gene ced-7 encodes a protein similar to ABC transporters. Cell. 1998;93:951–60. [PubMed: 9635425]
- 35.
- Moynault A , Luciani M F , Chimini G . ABC1, the mammalian homologue of the engulfment gene ced-7, is required during phagocytosis of both necrotic and apoptotic cells. Biochem Soc Trans. 1998;26:629–35. [PubMed: 10047795]
- 36.
- Langmann T , Klucken J , Reil M , Liebisch G , Luciani M F , Chimini G , Kaminski W E , Schmitz G . Molecular cloning of the human ATP-binding cassette transporter 1 (hABC1): evidence for sterol-dependent regulation in macrophages. Biochem Biophys Res Commun. 1999;257:29–33. [PubMed: 10092505]
- 37.
- Repa J J , Liang G , Ou J , Bashmakov Y , Lobaccaro J M , Shimomura I , Shan B , Brown M S , Goldstein J L , Mangelsdorf D J . Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRa and LXRb. Genes Dev. 2000;14:2819–30. [PMC free article: PMC317055] [PubMed: 11090130]
- 38.
- Santamarina-Fojo S , Peterson K , Knapper C , Qiu Y , Freeman L , Cheng J F , Osorio J , Remaley A , Yang X P , Haudenschild C , Prades C , Chimini G , Blackmon E , Francois T , Duverger N , Rubin E M , Rosier M , Denefle P , Fredrickson D S , Brewer H B Jr. Complete genomic sequence of the human ABCA1 gene: analysis of the human and mouse ATP-binding cassette A promoter. Proc Natl Acad Sci U S A. 2000;97:7987–7992. [PMC free article: PMC16657] [PubMed: 10884428]
- 39.
- Santamarina-Fojo S , Remaley A , Neufeld E , Brewer HB . Regulation and intracellular trafficking of ABCA1. J Lipid Res. 2001;42:1339–1345>. [PubMed: 11518753]
- 40.
- Pullinger C R , Hakamata H , Duchateau P N , Eng C , Aouizerat B E , Cho M H , Fielding C J , Kane J P . Analysis of hABC1 gene 5′ end: additional peptide sequence, promoter region, and four polymorphisms. Biochem Biophys Res Commun. 2000;271:451–5. [PubMed: 10799318]
- 41.
- Orso E , Broccardo C , Kaminski W E , Bottcher A , Liebisch G , Drobnik W , Gotz A , Chambenoit O , Diederich W , Langmann T , Spruss T , Luciani M F , Rothe G , Lackner K J , Chimini G , Schmitz G . Transport of lipids from Golgi to plasma membrane is defective in Tangier disease patients and Abc1-deficient mice. Nat Genet. 2000;24:192–6. [PubMed: 10655069]
- 42.
- McNeish J , Aiello R J , Guyot D , Turi T , Gabel C , Aldinger C , Hoppe K L , Roach M L , Royer L J , de Wet J , Broccardo C , Chimini G , Francone O L . High density lipoprotein deficiency and foam cell accumulation in mice with targeted disruption of ATP-binding cassette transporter-1. Proc Natl Acad Sci U S A. 2000;97:4245–50. [PMC free article: PMC18215] [PubMed: 10760292]
- 43.
- Groen A K , Bloks V W , Bandsma R H , Ottenhoff R , Chimini G , Kuipers F . Hepatobiliary cholesterol transport is not impaired in Abca1-null mice lacking HDL. J Clin Invest. 2001;108:843–50. [PMC free article: PMC200929] [PubMed: 11560953]
- 44.
- Joyce C W , Amar M J , Lambert G , Vaisman B L , Paigen B , Najib-Fruchart J , Hoyt R F Jr., Neufeld E D , Remaley A T , Fredrickson D S , Brewer H B Jr., Santamarina-Fojo S . The ATP binding cassette transporter A1 (ABCA1) modulates the development of aortic atherosclerosis in C57BL/6 and apoE-knockout mice. Proc Natl Acad Sci U S A. 2002;99:407–12. [PMC free article: PMC117573] [PubMed: 11752403]
- 45.
- Singaraja R R , Fievet C , Castro G , James E R , Hennuyer N , Clee S M , Bissada N , Choy J C , Fruchart J C , McManus B M , Staels B , Hayden M R . Increased ABCA1 activity protects against atherosclerosis. J Clin Invest. 2002;110:35–42. [PMC free article: PMC151034] [PubMed: 12093886]
- 46.
- Schreyer S A , Hart L K , Attie A D . Hypercatabolism of lipoprotein-free apolipoprotein A-I in HDL-deficient mutant chickens. Arterioscler Thromb. 1994;14:2053–9. [PubMed: 7981195]
- 47.
- Attie A D , Kastelein J P , Hayden M R . Pivotal role of ABCA1 in reverse cholesterol transport influencing HDL levels and susceptibility to atherosclerosis. J Lipid Res. 2001;42:1717–26. [PubMed: 11714841]
- 48.
- Allikmets R , Gerrard B , Glavac D , Ravnik-Glavac M , Jenkins N A , Gilbert D J , Copeland N G , Modi W , Dean M . Characterization and mapping of three new mammalian ATP-binding transporter genes from an EST database. Mamm Genome. 1995;6:114–7. [PubMed: 7766993]
- 49.
- Laing N M , Belinsky M G , Kruh G D , Bell D W , Boyd J T , Barone L , Testa J R , Tew K D . Amplification of the ATP-binding cassette 2 transporter gene is functionally linked with enhanced efflux of estramustine in ovarian carcinoma cells. Cancer Res. 1998;58:1332–7. [PubMed: 9537224]
- 50.
- Kaminski W E , Piehler A , Pullmann K , Porsch-Ozcurumez M , Duong C , Bared G M , Buchler C , Schmitz G . Complete coding sequence, promoter region, and genomic structure of the human ABCA2 gene and evidence for sterol-dependent regulation in macrophages. Biochem Biophys Res Commun. 2001;281:249–58. [PubMed: 11178988]
- 51.
- Zhou C , Zhao L , Nagaki N , Guan J , Nakajo S , Hirabayashi T , Kikuyama S , Shioda S . ATP-binding cassette transporter ABC2/ABCA2 in the rat brain: a novel mammalian lysosome-associated membrane protein and a specific marker for oligodendrocytes but not for myelin sheaths. J Neurosci. 2001;21:849–57. [PMC free article: PMC6762300] [PubMed: 11157071]
- 52.
- Allikmets R , Gerrard B , Hutchinson A , Dean M . Characterization of the human ABC superfamily: isolation and mapping of 21 new genes using the expressed sequence tags database. Hum Mol Genet. 1996;5:1649–1655. [PubMed: 8894702]
- 53.
- Klugbauer N , Hofmann F . Primary structure of a novel ABC transporter with a chromosomal localization on the band encoding the multidrug resistance-associated protein. FEBS Lett. 1996;391:61–5. [PubMed: 8706931]
- 54.
- Mulugeta S , Gray J M , Notarfrancesco K L , Gonzales L W , Koval M , Feinstein S I , Ballard P L , Fisher A B , Shuman H . Identification of LBM180, a lamellar body limiting membrane protein of alveolar type II cells, as the ABC transporter protein ABCA3. J Biol Chem. 2002;277:22147–55. [PubMed: 11940594]
- 55.
- Zen K , Notarfrancesco K , Oorschot V , Slot J W , Fisher A B , Shuman H . Generation and characterization of monoclonal antibodies to alveolar type II cell lamellar body membrane. Am J Physiol. 1998;275:L172–83. [PubMed: 9688949]
- 56.
- Yamano G , Funahashi H , Kawanami O , Zhao L X , Ban N , Uchida Y , Morohoshi T , Ogawa J , Shioda S , Inagaki N . ABCA3 is a lamellar body membrane protein in human lung alveolar type II cells. FEBS Lett. 2001;508:221–5. [PubMed: 11718719]
- 57.
- Allikmets R , Singh N , Sun H , Shroyer N F , Hutchinson A , Chidambaram A , Gerrard B , Baird L , Stauffer D , Peiffer A , Rattner A , Smallwood P , Li Y , Anderson K L , Lewis R A , Nathans J , Leppert M , Dean M , Lupski J R . A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet. 1997;15:236–246. [PubMed: 9054934]
- 58.
- Azarian S M , Travis G H . The photoreceptor rim protein is an ABC transporter encoded by the gene for recessive Stargardt's disease (ABCR). FEBS Lett. 1997;409:247–252. [PubMed: 9202155]
- 59.
- Sun H , Molday R S , Nathans J . Retinal stimulates ATP hydrolysis by purified and reconstituted ABCR, the photoreceptor-specific ATP-binding cassette transporter responsible for Stargardt disease. J Biol Chem. 1999;274:8269–81. [PubMed: 10075733]
- 60.
- Weng J , Mata N L , Azarian S M , Tzekov R T , Birch D G , Travis G H . Insights into the function of Rim protein in photoreceptors and etiology of Stargardt's disease from the phenotype in abcr knockout mice. Cell. 1999;98:13–23. [PubMed: 10412977]
- 61.
- Allikmets R . Simple and complex ABCR: genetic predisposition to retinal disease. Am J Hum Genet. 2000;67:793–799. [PMC free article: PMC1287884] [PubMed: 10970771]
- 62.
- Martinez-Mir A , Paloma E , Allikmets R , Ayuso C , del Rio T , Dean M , Vilageliu L , Gonzalez-Duarte R , Balcells S . Retinitis pigmentosa caused by a homozygous mutation in the Stargardt disease gene ABCR [letter; comment] Nat Genet. 1998;18:11–2. [PubMed: 9425888]
- 63.
- Rozet J M , Gerber S , Ghazi I , Perrault I , Ducroq D , Souied E , Cabot A , Dufier J L , Munnich A , Kaplan J . Mutations of the retinal specific ATP binding transporter gene (ABCR) in a single family segregating both autosomal recessive retinitis pigmentosa RP19 and Stargardt disease: evidence of clinical heterogeneity at this locus. J Med Genet. 1999;36:447–51. [PMC free article: PMC1734380] [PubMed: 10874631]
- 64.
- Cremers F P , van de Pol D J , van Driel M , den Hollander A I , van Haren F J , Knoers N V , Tijmes N , Bergen A A , Rohrschneider K , Blankenagel A , Pinckers A J , Deutman A F , Hoyng C B . Autosomal recessive retinitis pigmentosa and cone-rod dystrophy caused by splice site mutations in the Stargardt's disease gene ABCR. Hum Mol Genet. 1998;7:355–362. [PubMed: 9466990]
- 65.
- Stargardt K . Uber familiare, progressive degeneration in der maculagegend des auges. Albrecht van Graefes Arch. Ophthalmol. 1909;71:534–550.
- 66.
- Allikmets R , Shroyer N F , Singh N , Seddon J M , Lewis R A , Bernstein P S , Peiffer A , Zabriskie N A , Li Y , Hutchinson A , Dean M , Lupski J R , Leppert M . Mutation of the Stargardt disease gene (ABCR) in age-related macular degeneration. Science. 1997;277:1805–1807. [PubMed: 9295268]
- 67.
- Mata N L , Tzekov R T , Liu X , Weng J , Birch D G , Travis G H . Delayed dark-adaptation and lipofuscin accumulation in abcr +/– mice: implications for involvement of ABCR in age-related macular degeneration. Invest Ophthalmol Vis Sci. 2001;42:1685–90. [PubMed: 11431429]
- 68.
- Schriml L M , Dean M . Identification of 18 mouse ABC genes and characterization of the ABC superfamily in Mus musculus. Genomics. 2000;64:24–31. [PubMed: 10708515]
- 69.
- Kaminski W E , Wenzel J J , Piehler A , Langmann T , Schmitz G . ABCA6, a novel a subclass ABC transporter. Biochem Biophys Res Commun. 2001;285:1295–301. [PubMed: 11478798]
- 70.
- Broccardo C , Osorio J , Luciani M F , Schriml L , Prades C , Shulenin S , Arnould I , Naudin L , Lafarge C , Rosier M , Jordan B , Mattei M G , Dean M , Denefle P , Chimini G . Comparative analysis of promoter structure and genomic organization of human and mouse ABCA7, a novel ABCA transporter. Biochim Biophys Res Commun. 2001;92:264–270. [PubMed: 11435699]
- 71.
- Kaminski W E , Orso E , Diederich W , Klucken J , Drobnik W , Schmitz G . Identification of a novel human sterol-sensitive ATP-binding cassette transporter (ABCA7). Biochem Biophys Res Commun. 2000;273:532–8. [PubMed: 10873640]
- 72.
- Kaminski W E , Piehler A , Schmitz G . Genomic organization of the human cholesterol-responsive ABC transporter ABCA7: tandem linkage with the minor histocompatibility antigen HA-1 gene. Biochem Biophys Res Commun. 2000;278:782–9. [PubMed: 11095984]
- 73.
- Tanaka A R , Keda Y , Abe-Dohmae S , Arakawa R , Sadanami K , Kidera A , Nakagawa S , Nagase T , Aoki R , Kioka N , Amachi T , Yokoyama S , Ueda K . Human ABCA1 contains a large amino-terminal extracellular domain homologous to an epitope of Sjogren's Syndrome. Biochem Biophys Res Commun. 2001;283:1019–25. [PubMed: 11355874]
- 74.
- Iida A , Saito S , Sekine A , Mishima C , Kitamura Y , Kondo K , Harigae S , Osawa S , Nakamura Y . Catalog of 605 single-nucleotide polymorphisms (SNPs) among 13 genes encoding human ATP-binding cassette transporters: ABCA4, ABCA7, ABCA8, ABCD1, ABCD3, ABCD4, ABCE1, ABCF1, ABCG1, ABCG2, ABCG4, ABCG5, and ABCG8. J Hum Genet. 2002;47:285–310. [PubMed: 12111378]
- 75.
- Piehler A , Kaminski W E , Wenzel J J , Langmann T , Schmitz G . Molecular structure of a novel cholesterol-responsive A subclass ABC transporter, ABCA9. Biochem Biophys Res Commun. 2002;295:408–16. [PubMed: 12150964]
- 76.
- Juliano R L , Ling V A . A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta. 1976;455:152–162. [PubMed: 990323]
- 77.
- Riordan J R , Deuchars K , Kartner N , Alon N , Trent J , Ling V . Amplification of P-glycoprotein genes in multidrug-resistant mammalian cell lines. Nature. 1985;316:817–9. [PubMed: 2863759]
- 78.
- Kartner N , Evernden-Porelle D , Bradley G , Ling V . Detection of P-glycoprotein in multidrug-resistant cell lines by monoclonal antibodies. Nature. 1985;316:820–3. [PubMed: 2412130]
- 79.
- Roninson I B , Chin J E , Choi K , Gros P , Housman D E , Fojo A , Shen D -W , Gottesman M M , Pastan I . Isolation of human mdr DNA sequences amplified in multidrug-resistant KB carcinoma cells. Proc Natl Acad Sci U S A. 1986;83:4538–4542. [PMC free article: PMC323769] [PubMed: 3459187]
- 80.
- Ambudkar S V , Gottesman M M . ABC transporters: biochemical, cellular, and molecular aspects. Methods Enzymol. 1998;292:1–853.
- 81.
- Hoffmeyer S , Burk O , von Richter O , Arnold H P , Brockmoller J , Johne A , Cascorbi I , Gerloff T , Roots I , Eichelbaum M , Brinkmann U . Functional polymorphisms of the human multidrug-resistance gene: multiple sequence variations and correlation of one allele with P-glycoprotein expression and activity in vivo. Proc Natl Acad Sci U S A. 2000;97:3473–8. [PMC free article: PMC16264] [PubMed: 10716719]
- 82.
- Schinkel A H , Smit J J , van Tellingen O , Beijnen J H , Wagenaar E , van Deemter L , Mol C A , van der Valk M A , Robanus-Maandag E C , te Riele H P . et al. Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood–brain barrier and to increased sensitivity to drugs. Cell. 1994;77:491–502. [PubMed: 7910522]
- 83.
- Schinkel A H , Mayer U , Wagenaar E , Mol C A , van Deemter L , Smit J J , van der Valk M A , Voordouw A C , Spits H , van Tellingen O , Zijlmans J M , Fibbe W E , Borst P . Normal viability and altered pharmacokinetics in mice lacking mdr1-type (drug-transporting) P-glycoproteins. Proc Natl Acad Sci U S A. 1997;94:4028–33. [PMC free article: PMC20562] [PubMed: 9108099]
- 84.
- Mealey K L , Bentjen S A , Gay J M , Cantor G H . Ivermectin sensitivity in collies is associated with a deletion mutation of the mdr1 gene. Pharmacogenetics. 2001;11:727–33. [PubMed: 11692082]
- 85.
- Chaudhary P M , Roninson I B . Expression and activity of P-glycoprotein, a multidrug efflux pump, in human hematopoietic stem cells. Cell. 1991;66:85–94. [PubMed: 1712673]
- 86.
- Randolph G J , Beaulieu S , Pope M , Sugawara I , Hoffman L , Steinman R M , Muller W A . A physiologic function for P-glycoprotein (MDR-1) during the migration of dendritic cells from skin via afferent lymphatic vessels. Proc Natl Acad Sci U S A. 1998;95:6924–9. [PMC free article: PMC22688] [PubMed: 9618515]
- 87.
- Trowsdale J , Hanson I , Mockridge I , Beck S , Townsend A , Kelly A . Sequences encoded in the class II region of the MHC related to the “ABC” superfamily of transporters [see comments] Nature. 1990;348:741–4. [PubMed: 2259383]
- 88.
- Spies T , Bresnahan M , Bahram S , Arnold D , Blanck G , Mellins E , Pious D , DeMars R . A gene in the human major histocompatibility complex class II region controlling the class I antigen presentation pathway [see comments] Nature. 1990;348:744–7. [PubMed: 2259384]
- 89.
- Monaco J J , Cho S , Attaya M . Transport protein genes in the murine MHC: possible implications for antigen processing. Science. 1990;250:1723–6. [PubMed: 2270487]
- 90.
- Salter R D , Cresswell P . Impaired assembly and transport of HLA-A and -B antigens in a mutant TxB cell hybrid. EMBO J. 1986;5:943–9. [PMC free article: PMC1166886] [PubMed: 3522223]
- 91.
- Shepherd J C , Schumacher T N , Ashton-Rickardt P G , Imaeda S , Ploegh H L , Janeway C A Jr, Tonegawa S . TAP1-dependent peptide translocation in vitro is ATP dependent and peptide selective. Cell. 1993;74:577–84. [PubMed: 8348620]
- 92.
- Androlewicz M J , Anderson K S , Cresswell P . Evidence that transporters associated with antigen processing translocate a major histocompatibility complex class I-binding peptide into the endoplasmic reticulum in an ATP-dependent manner. Proc Natl Acad Sci U S A. 1993;90:9130–4. [PMC free article: PMC47515] [PubMed: 8415666]
- 93.
- Uebel S , Tampe R . Specificity of the proteasome and the TAP transporter. Curr Opin Immunol. 1999;11:203–8. [PubMed: 10322157]
- 94.
- Uebel S , Kraas W , Kienle S , Wiesmuller K H , Jung G , Tampe R . Recognition principle of the TAP transporter disclosed by combinatorial peptide libraries. Proc Natl Acad Sci U S A. 1997;94:8976–81. [PMC free article: PMC22991] [PubMed: 9256420]
- 95.
- Van Kaer L , Ashton-Rickardt P G , Ploegh H L , Tonegawa S . TAP1 mutant mice are deficient in antigen presentation, surface class I molecules, and CD4-8+ T cells. Cell. 1992;71:1205–14. [PubMed: 1473153]
- 96.
- Lacaille V G , Androlewicz M J . Herpes simplex virus inhibitor ICP47 destabilizes the transporter associated with antigen processing (TAP) heterodimer. J Biol Chem. 1998;273:17386–90. [PubMed: 9651323]
- 97.
- Galocha B , Hill A , Barnett B C , Dolan A , Raimondi A , Cook R F , Brunner J , McGeoch D J , Ploegh H L . The active site of ICP47, a herpes simplex virus-encoded inhibitor of the major histocompatibility complex (MHC)-encoded peptide transporter associated with antigen processing (TAP), maps to the NH2-terminal 35 residues. J Exp Med. 1997;185:1565–72. [PMC free article: PMC2196299] [PubMed: 9151894]
- 98.
- York I A , Roop C , Andrews D W , Riddell S R , Graham F L , Johnson D C . A cytosolic herpes simplex virus protein inhibits antigen presentation to CD8+ T lymphocytes. Cell. 1994;77:525–35. [PubMed: 8187174]
- 99.
- Hill A B , Barnett B C , McMichael A J , McGeoch D J . HLA class I molecules are not transported to the cell surface in cells infected with herpes simplex virus types 1 and 2. J Immunol. 1994;152:2736–41. [PubMed: 8144880]
- 100.
- Chen H L , Gabrilovich D , Tampe R , Girgis K R , Nadaf S , Carbone D P . A functionally defective allele of TAP1 results in loss of MHC class I antigen presentation in a human lung cancer. Nat Genet. 1996;13:210–3. [PubMed: 8640228]
- 101.
- de la Salle H , Zimmer J , Fricker D , Angenieux C , Cazenave J P , Okubo M , Maeda H , Plebani A , Tongio M M , Dormoy A , Hanau D . HLA class I deficiencies due to mutations in subunit 1 of the peptide transporter TAP1. J Clin Invest. 1999;103:R9–R13. [PMC free article: PMC408129] [PubMed: 10074495]
- 102.
- de la Salle H , Hanau D , Fricker D , Urlacher A , Kelly A , Salamero J , Powis S H , Donato L , Bausinger H , Laforet M . et al. Homozygous human TAP peptide transporter mutation in HLA class I deficiency. Science. 1994;265:237–41. [PubMed: 7517574]
- 103.
- Smit J J , Schinkel A H , Oude Elferink R P , Groen A K , Wagenaar E , van Deemter L , Mol C A , Ottenhoff R , van der Lugt N M , van Roon M A . et al. Homozygous disruption of the murine mdr2 P-glycoprotein gene leads to a complete absence of phospholipid from bile and to liver disease. Cell. 1993;75:451–62. [PubMed: 8106172]
- 104.
- Ruetz S , Gros P . Phosphatidylcholine translocase: a physiological role for the mdr2 gene. Cell. 1994;77:1071–81. [PubMed: 7912658]
- 105.
- van Helvoort A , Smith A J , Sprong H , Fritzsche I , Schinkel A H , Borst P , van Meer G . MDR1 P-glycoprotein is a lipid translocase of broad specificity, while MDR3 P-glycoprotein specifically translocates phosphatidylcholine. Cell. 1996;87:507–17. [PubMed: 8898203]
- 106.
- Deleuze J F , Jacquemin E , Dubuisson C , Cresteil D , Dumont M , Erlinger S , Bernard O , Hadchouel M . Defect of multidrug-resistance 3 gene expression in a subtype of progressive familial intrahepatic cholestasis. Hepatology. 1996;23:904–8. [PubMed: 8666348]
- 107.
- de Vree J M , Jacquemin E , Sturm E , Cresteil D , Bosma P J , Aten J , Deleuze J F , Desrochers M , Burdelski M , Bernard O , Oude Elferink R P , Hadchouel M . Mutations in the MDR3 gene cause progressive familial intrahepatic cholestasis. Proc Natl Acad Sci U S A. 1998;95:282–7. [PMC free article: PMC18201] [PubMed: 9419367]
- 108.
- Dixon P H , Weerasekera N , Linton K J , Donaldson O , Chambers J , Egginton E , Weaver J , Nelson-Piercy C , de Swiet M , Warnes G , Elias E , Higgins C F , Johnston D G , McCarthy M I , Williamson C . Heterozygous MDR3 missense mutation associated with intrahepatic cholestasis of pregnancy: evidence for a defect in protein trafficking. Hum Mol Genet. 2000;9:1209–17. [PubMed: 10767346]
- 109.
- Jacquemin E , Cresteil D , Manouvrier S , Boute O , Hadchouel M . Heterozygous non-sense mutation of the MDR3 gene in familial intrahepatic cholestasis of pregnancy. Lancet. 1999;353:210–1. [PubMed: 9923886]
- 110.
- Mitsuhashi N , Miki T , Senbongi H , Yokoi N , Yano H , Miyazaki M , Nakajima N , Wanaga T , Yokoyama Y , Shibata T , Seino S . MTABC3, a novel mitochondrial ATP-binding cassette protein involved in iron homeostasis. J Biol Chem. 2000;275:17536–40. [PubMed: 10837493]
- 111.
- Csere P , Lill R , Kispal G . Identification of a human mitochondrial ABC transporter, the functional orthologue of yeast Atm1p. FEBS Lett. 1998;441:266–70. [PubMed: 9883897]
- 112.
- Allikmets R , Raskind W H , Hutchinson A , Schueck N D , Dean M , Koeller D M . Mutation of a putative mitochondrial iron transporter gene (ABC7) in X- linked sideroblastic anemia and ataxia (XLSA/A). Hum Mol Genet. 1999;8:743–9. [PubMed: 10196363]
- 113.
- Bekri S , Kispal G , Lange H , Fitzsimons E , Tolmie J , Lill R , Bishop D F . Human ABC7 transporter: gene structure and mutation causing X-linked sideroblastic anemia with ataxia with disruption of cytosolic iron- sulfur protein maturation. Blood. 2000;96:3256–64. [PubMed: 11050011]
- 114.
- Hogue D L , Liu L , Ling V . Identification and characterization of a mammalian mitochondrial ATP- binding cassette membrane protein. J Mol Biol. 1999;285:379–89. [PubMed: 9878413]
- 115.
- Zhang F , Zhang W , Liu L , Fisher C L , Hui D , Childs S , Dorovini-Zis K , Ling V . Characterization of ABCB9, an ATP binding cassette protein associated with lysosomes. J Biol Chem. 2000;275:23287–94. [PubMed: 10748049]
- 116.
- Zhang F , Hogue D L , Liu L , Fisher C L , Hui D , Childs S , Ling V . M-ABC2, a new human mitochondrial ATP-binding cassette membrane protein. FEBS Lett. 2000;478:89–94. [PubMed: 10922475]
- 117.
- Childs S , Yeh R L , Georges E , Ling V . Identification of a sister gene to P-glycoprotein. Cancer Res. 1995;55:2029–34. [PubMed: 7538046]
- 118.
- Gerloff T , Stieger B , Hagenbuch B , Madon J , Landmann L , Roth J , Hofmann A F , Meier P J . The sister of P-glycoprotein represents the canalicular bile salt export pump of mammalian liver. J Biol Chem. 1998;273:10046–50. [PubMed: 9545351]
- 119.
- Strautnieks S , Bull L N , Knisely A S , Kocoshis S A , Dahl N , Arnell H , Sokal E , Dahan K , Childs S , Ling V , Tanner M S , Kagalwalla A F , Nemeth A , Pawlowska J , Baker A , Mieli-Vergani G , Freimer N B , Gardiner R M , Thompson R J . A gene encoding a liver-specific ABC transporter is mutated in progressive familial intrahepatic cholestasis. Nat Genet. 1998;20:233–238. [PubMed: 9806540]
- 120.
- Wang R , Salem M , Yousef I M , Tuchweber B , Lam P , Childs S J , Helgason C D , Ackerley C , Phillips M J , Ling V . Targeted inactivation of sister of P-glycoprotein gene (spgp) in mice results in nonprogressive but persistent intrahepatic cholestasis. Proc Natl Acad Sci U S A. 2001;98:2011–6. [PMC free article: PMC29373] [PubMed: 11172067]
- 121.
- Plass J R , Mol O , Heegsma J , Geuken M , Faber K N , Jansen P L , Muller M . Farnesoid X receptor and bile salts are involved in transcriptional regulation of the gene encoding the human bile salt export pump. Hepatology. 2002;35:589–96. [PubMed: 11870371]
- 122.
- Cole S P , Deeley R G . Multidrug resistance mediated by the ATP-binding cassette transporter protein MRP. Bioessays. 1998;20:931–40. [PubMed: 9872059]
- 123.
- Cole S P C , Bhardwaj G , Gerlach J H , Mackie J E , Grant C E , Almquist K C , Stewart A J , Kurz E U , Duncan A M V , Deeley R G . Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science. 1992;258:1650–1654. [PubMed: 1360704]
- 124.
- Kuwano M , Toh S , Uchiumi T , Takano H , Kohno K , Wada M . Multidrug resistance-associated protein subfamily transporters and drug resistance. Anticancer Drug Des. 1999;14:123–31. [PubMed: 10405639]
- 125.
- Zaman G J , Flens M J , van Leusden M R , de Haas M , Mulder H S , Lankelma J , Pinedo H M , Scheper R J , Baas F , Broxterman H J . et al. The human multidrug resistance-associated protein MRP is a plasma membrane drug-efflux pump. Proc Natl Acad Sci U S A. 1994;91:8822–8826. [PMC free article: PMC44698] [PubMed: 7916458]
- 126.
- Cole S P , Sparks K E , Fraser K , Loe D W , Grant C E , Wilson G M , Deeley R G . Pharmacological characterization of multidrug resistant MRP-transfected human tumor cells. Cancer Res. 1994;54:5902–10. [PubMed: 7954421]
- 127.
- Leier I , Jedlitschky G , Buchholz U , Cole S P , Deeley R G , Keppler D . The MRP gene encodes an ATP-dependent export pump for leukotriene C4 and structurally related conjugates. J Biol Chem. 1994;269:27807–10. [PubMed: 7961706]
- 128.
- Lorico A , Rappa G , Finch R A , Yang D , Flavell R A , Sartorelli A C . Disruption of the murine MRP (multidrug resistance protein) gene leads to increased sensitivity to etoposide (VP-16) and increased levels of glutathione. Cancer Res. 1997;57:5238–42. [PubMed: 9393741]
- 129.
- Wijnholds J , Evers R , van Leusden M R , Mol C A , Zaman G J , Mayer U , Beijnen J H , van der Valk M , Krimpenfort P , Borst P . Increased sensitivity to anticancer drugs and decreased inflammatory response in mice lacking the multidrug resistance-associated protein. Nat Med. 1997;3:1275–9. [PubMed: 9359705]
- 130.
- Robbiani D F , Finch R A , Jager D , Muller W A , Sartorelli A C , Randolph G J . The leukotriene C(4) transporter MRP1 regulates CCL19 (MIP-3β, ELC)-dependent mobilization of dendritic cells to lymph nodes. Cell. 2000;103:757–68. [PubMed: 11114332]
- 131.
- Kool M , de Haas M , Scheffer G L , Scheper R J , van Eijk M J , Juijn J A , Baas F , Borst P . Analysis of expression of cMOAT (MRP2), MRP3, MRP4, and MRP5, homologues of the multidrug resistance-associated protein gene (MRP1), in human cancer cell lines. Cancer Res. 1997;57:3537–47. [PubMed: 9270026]
- 132.
- Paulusma C C , Bosma P J , Zaman G J R , Bakker C T M , Otter M , Scheffer G L , Scheper R J , Borst P , Oude Elferink R P J . Congenital jaundice in rats with a mutation in a multidrug resistance-associated protein gene. Science. 1996;271:1126–1128. [PubMed: 8599091]
- 133.
- Wada M , Toh S , Taniguchi K , Nakamura T , Uchiumi T , Kohno K , Yoshida I , Kimura A , Sakisaka S , Adachi Y , Kuwano M . Mutations in the canilicular multispecific organic anion transporter (cMOAT) gene, a novel ABC transporter, in patients with hyperbilirubinemia II/Dubin-Johnson syndrome. Hum Mol Genet. 1998;7:203–7. [PubMed: 9425227]
- 134.
- Toh S , Wada M , Uchiumi T , nokuchi A , Makino Y , Horie Y , Adachi Y , Sakisaka S , Kuwano M . Genomic structure of the canalicular multispecific organic anion- transporter gene (MRP2/cMOAT) and mutations in the ATP-binding-cassette region in Dubin-Johnson syndrome. Am J Hum Genet. 1999;64:739–46. [PMC free article: PMC1377791] [PubMed: 10053008]
- 135.
- Cui Y , Konig J , Buchholz J K , Spring H , Leier I , Keppler D . Drug resistance and ATP-dependent conjugate transport mediated by the apical multidrug resistance protein, MRP2, permanently expressed in human and canine cells. Mol Pharmacol. 1999;55:929–37. [PubMed: 10220572]
- 136.
- Kikuchi S , Hata M , Fukumoto K , Yamane Y , Matsui T , Tamura A , Yonemura S , Yamagishi H , Keppler D , Tsukita S . Radixin deficiency causes conjugated hyperbilirubinemia with loss of Mrp2 from bile canalicular membranes. Nat Genet. 2002;31:320–5. [PubMed: 12068294]
- 137.
- Zeng H , Bain L J , Belinsky M G , Kruh G D . Expression of multidrug resistance protein-3 (multispecific organic anion transporter-D) in human embryonic kidney 293 cells confers resistance to anticancer agents. Cancer Res. 1999;59:5964–7. [PubMed: 10606242]
- 138.
- Kool M , van der Linden M , de Haas M , Scheffer G L , de Vree J M , Smith A J , Jansen G , Peters G J , Ponne N , Scheper R J , Elferink R P , Baas F , Borst P . MRP3, an organic anion transporter able to transport anti-cancer drugs. Proc Natl Acad Sci U S A. 1999;96:6914–9. [PMC free article: PMC22016] [PubMed: 10359813]
- 139.
- Lee K , Belinsky M G , Bell D W , Testa J R , Kruh G D . Isolation of MOAT-B, a widely expressed multidrug resistance-associated protein/canalicular multispecific organic anion transporter-related transporter. Cancer Res. 1998;58:2741–7. [PubMed: 9661885]
- 140.
- Schuetz J D , Connelly M C , Sun D , Paibir S G , Flynn P M , inivas R V , Kumar A , Fridland A . MRP4: a previously unidentified factor in resistance to nucleoside-based antiviral drugs. Nat Med. 1999;5:1048–51. [PubMed: 10470083]
- 141.
- Belinsky M G , Bain L J , Balsara B B , Testa J R , Kruh G D . Characterization of MOAT-C and MOAT-D, new members of the MRP/cMOAT subfamily of transporter proteins. J Natl Cancer Inst. 1998;90:1735–41. [PubMed: 9827529]
- 142.
- Wijnholds J , Mol C A , van Deemter L , de Haas M , Scheffer G L , Baas F , Beijnen J H , Scheper R J , Hatse S , De Clercq E , Balzarini J , Borst P . Multidrug-resistance protein 5 is a multispecific organic anion transporter able to transport nucleotide analogs. Proc Natl Acad Sci U S A. 2000;97:7476–81. [PMC free article: PMC16570] [PubMed: 10840050]
- 143.
- Bergen A A , Plomp A S , Schuurman E J , Terry S , Breuning M , Dauwerse H , Swart J , Kool M , van Soest S , Baas F , ten Brink J B , de Jong P T . Mutations in ABCC6 cause pseudoxanthoma elasticum. Nat Genet. 2000;25:228–31. [PubMed: 10835643]
- 144.
- Le Saux O , Urban Z , Tschuch C , Csiszar K , Bacchelli B , Quaglino D , Pasquali-Ronchetti I , Pope F M , Richards A , Terry S , Bercovitch L , de Paepe A , Boyd C D . Mutations in a gene encoding an ABC transporter cause pseudoxanthoma elasticum. Nat Genet. 2000;25:223–7. [PubMed: 10835642]
- 145.
- Le Saux O , Beck K , Sachsinger C , Silvestri C , Treiber C , Goring H H , Johnson E W , De Paepe A , Pope F M , Pasquali-Ronchetti I , Bercovitch L , Terry S , Boyd C D , Marais A S , Viljoen D L . A spectrum of ABCC6 mutations is responsible for pseudoxanthoma elasticum. Am J Hum Genet. 2001;69:749–64. [PMC free article: PMC1226061] [PubMed: 11536079]
- 146.
- Ringpfeil F , Lebwohl M G , Christiano A M , Uitto J . Pseudoxanthoma elasticum: mutations in the MRP6 gene encoding a transmembrane ATP-binding cassette (ABC) transporter. Proc Natl Acad Sci U S A. 2000;97:6001–6. [PMC free article: PMC18548] [PubMed: 10811882]
- 147.
- Struk B , Cai L , Zach S , Ji W , Chung J , Lumsden A , Stumm M , Huber M , Schaen L , Kim C A , Goldsmith L A , Viljoen D , Figuera L E , Fuchs W , Munier F , Ramesar R , Hohl D , Richards R , Neldner K H , Lindpaintner K . Mutations of the gene encoding the transmembrane transporter protein ABC-C6 cause pseudoxanthoma elasticum. J Mol Med. 2000;78:282–6. [PubMed: 10954200]
- 148.
- Wang J , Near S , Young K , Connelly P W , Hegele R A . ABCC6 gene polymorphism associated with variation in plasma lipoproteins. J Hum Genet. 2001;46:699–705. [PubMed: 11776382]
- 149.
- Pulkkinen L , Nakano A , Ringpfeil F , Uitto J . Identification of ABCC6 pseudogenes on human chromosome 16p: implications for mutation detection in pseudoxanthoma elasticum. Hum Genet. 2001;109:356–65. [PubMed: 11702217]
- 150.
- Ilias A , Urban Z , Seidl T L , Le Saux O , Sinko E , Boyd C D , Sarkadi B , Varadi A . Loss of ATP-dependent transport activity in pseudoxanthoma elasticum- associated mutants of human ABCC6 (MRP6). J Biol Chem. 2002;277:16860–7. [PubMed: 11880368]
- 151.
- Cai J , Daoud R , Alqawi O , Georges E , Pelletier J , Gros P . Nucleotide binding and nucleotide hydrolysis properties of the ABC transporter MRP6 (ABCC6). Biochemistry. 2002;41:8058–67. [PubMed: 12069597]
- 152.
- Madon J , Hagenbuch B , Landmann L , Meier P J , Stieger B . Transport function and hepatocellular localization of mrp6 in rat liver. Mol Pharmacol. 2000;57:634–41. [PubMed: 10692506]
- 153.
- Trip M D , Smulders Y M , Wegman J J , Hu X , Boer J M , ten Brink J B , Zwinderman A H , Kastelein J J , Feskens E J , Bergen A A . Frequent mutation in the ABCC6 gene (R1141X) is associated with a strong increase in the prevalence of coronary artery disease. Circulation. 2002;106:773–5. [PubMed: 12176944]
- 154.
- Riordan J R , Rommens J M , Kerem B , Alon N , Rozmahel R , Grzelczak Z , Zielenski J , Lok S , Plavsic N , Chou J L . et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science. 1989;245:1066–73. [PubMed: 2475911]
- 155.
- Kerem B , Rommens J M , Buchanan J A , Markiewicz D , Cox T K , Chakravarti A , Buchwald M , Tsui L C . Identification of the cystic fibrosis gene: genetic analysis. Science. 1989;245:1073–80. [PubMed: 2570460]
- 156.
- Rommens J M , Iannuzzi M C , Kerem B , Drumm M L , Melmer G , Dean M , Rozmahel R , Cole J L , Kennedy D , Hidaka N . et al. Identification of the cystic fibrosis gene: chromosome walking and jumping. Science. 1989;245:1059–65. [PubMed: 2772657]
- 157.
- Andersen D H . Cystic fibrosis of the pancreas and its relationship to celiac disease. Amer J Dis Child. 1938;56:344–399.
- 158.
- Estivill X , Bancells C , Ramos C . Geographic distribution and regional origin of 272 cystic fibrosis mutations in European populations. The Biomed CF Mutation Analysis Consortium. Hum Mutat. 1997;10:135–54. [PubMed: 9259197]
- 159.
- Quinton P M . Human genetics. What is good about cystic fibrosis? Curr Biol. 1994;4:742–3. [PubMed: 7525005]
- 160.
- Gabriel S E , Clarke L L , Boucher R C , Stutts M J . CFTR and outward rectifying chloride channels are distinct proteins with a regulatory relationship. Nature. 1993;363:263–8. [PubMed: 7683773]
- 161.
- Pier G B , Grout M , Zaidi T , Meluleni G , Mueschenborn S S , Banting G , Ratcliff R , Evans M J , Colledge W H . Salmonella typhi uses CFTR to enter intestinal epithelial cells. Nature. 1998;393:79–82. [PubMed: 9590693]
- 162.
- Dorin J R , Dickinson P , Alton E W , Smith S N , Geddes D M , Stevenson B J , Kimber W L , Fleming S , Clarke A R , Hooper M L . et al. Cystic fibrosis in the mouse by targeted insertional mutagenesis. Nature. 1992;359:211–5. [PubMed: 1382232]
- 163.
- Snouwaert J N , Brigman K K , Latour A M , Malouf N N , Boucher R C , Smithies O , Koller B H . An animal model for cystic fibrosis made by gene targeting. Science. 1992;257:1083–8. [PubMed: 1380723]
- 164.
- van Doorninck J H , French P J , Verbeek E , Peters R H , Morreau H , Bijman J , Scholte B J . A mouse model for the cystic fibrosis delta F508 mutation. EMBO J. 1995;14:4403–11. [PMC free article: PMC394531] [PubMed: 7556083]
- 165.
- Dean M , Santis G . Heterogeneity in the severity of cystic fibrosis and the role of CFTR gene mutations. Hum Genet. 1994;93:364–8. [PubMed: 7513291]
- 166.
- Tsui L C . The cystic fibrosis transmembrane conductance regulator gene. Am J Respir Crit Care Med. 1995;151:S47–53. [PubMed: 7533605]
- 167.
- Dean M , White M B , Amos J , Gerrard B , Stewart C , Khaw K T , Leppert M . Multiple mutations in highly conserved residues are found in mildly affected cystic fibrosis patients. Cell. 1990;61:863–70. [PubMed: 2344617]
- 168.
- Anguiano A , Oates R D , Amos J A , Dean M , Gerrard B , Stewart C , Maher T A , White M B , Milunsky A . Congenital bilateral absence of the vas deferens. A primarily genital form of cystic fibrosis. JAMA. 1992;267:1794–7. [PubMed: 1545465]
- 169.
- Dumur V , Gervais R , Rigot J M , Delomel-Vinner E , Decaestecker B , Lafitte J J , Roussel P . Congenital bilateral absence of the vas deferens (CBAVD) and cystic fibrosis transmembrane regulator (CFTR): correlation between genotype and phenotype. Hum Genet. 1996;97:7–10. [PubMed: 8557264]
- 170.
- Cohn J A , Friedman K J , Noone P G , Knowles M R , Silverman L M , Jowell P S . Relation between mutations of the cystic fibrosis gene and idiopathic pancreatitis. N Engl J Med. 1998;339:653–8. [PubMed: 9725922]
- 171.
- Pignatti P F , Bombieri C , Marigo C , Benetazzo M , Luisetti M . Increased incidence of cystic fibrosis gene mutations in adults with disseminated bronchiectasis. Hum Mol Genet. 1995;4:635–9. [PubMed: 7543317]
- 172.
- O'Dea S , Harrison D J . CFTR gene transfer to lung epithelium—on the trail of a target cell. Curr Gene Ther. 2002;2:173–81. [PubMed: 12109214]
- 173.
- Boucher R C . Status of gene therapy for cystic fibrosis lung disease. J Clin Invest. 1999;103:441–5. [PMC free article: PMC408111] [PubMed: 10021450]
- 174.
- Wine J J . The genesis of cystic fibrosis lung disease. J Clin Invest. 1999;103:309–12. [PMC free article: PMC407906] [PubMed: 9927490]
- 175.
- Wine J J , Brayden D J , Hagiwara G , Krouse M E , Law T C , Muller U J , Solc C K , Ward C L , Widdicombe J H , Xia Y . Cystic fibrosis, the CFTR, and rectifying Cl– channels. Adv Exp Med Biol. 1991;290:253–69. [PubMed: 1719763]
- 176.
- Riordan J R . The cystic fibrosis transmembrane conductance regulator. Annu Rev Physiol. 1993;55:609–30. [PubMed: 7682047]
- 177.
- Riordan J R . Cystic fibrosis as a disease of misprocessing of the cystic fibrosis transmembrane conductance regulator glycoprotein. Am J Hum Genet. 1999;64:1499–504. [PMC free article: PMC1377893] [PubMed: 10330337]
- 178.
- Ostedgaard L S , Baldursson O , Welsh M J . Regulation of the cystic fibrosis transmembrane conductance regulator Cl– channel by its R domain. J Biol Chem. 2001;276:7689–92. [PubMed: 11244086]
- 179.
- Thomas P M , Cote G J , Wohllk N , Haddad B , Mathew P M , Rabl W , Aguilar-Bryan L , Gagel R F , Bryan J . Mutations in the sulfonylurea receptor gene in familial persistent hyperinsulinemic hypoglycemia of infancy. Science. 1995;268:426–9. [PubMed: 7716548]
- 180.
- Seghers V , Nakazaki M , DeMayo F , Aguilar-Bryan L , Bryan J . Sur1 knockout mice. A model for K(ATP) channel-independent regulation of insulin secretion. J Biol Chem. 2000;275:9270–7. [PubMed: 10734066]
- 181.
- Goksel D L , Fischbach K , Duggirala R , Mitchell B D , Aguilar-Bryan L , Blangero J , Stern M P , O'Connell P . Variant in sulfonylurea receptor-1 gene is associated with high insulin concentrations in non-diabetic Mexican Americans: SUR-1 gene variant and hyperinsulinemia. Hum Genet. 1998;103:280–5. [PubMed: 9799081]
- 182.
- Reis A F , Ye W Z , Dubois-Laforgue D , Bellanne-Chantelot C , Timsit J , Velho G . Association of a variant in exon 31 of the sulfonylurea receptor 1 (SUR1) gene with type 2 diabetes mellitus in French Caucasians. Hum Genet. 2000;107:138–44. [PubMed: 11030411]
- 183.
- Altshuler D , Hirschhorn J N , Klannemark M , Lindgren C M , Vohl M C , Nemesh J , Lane C R , Schaffner S F , Bolk S , Brewer C , Tuomi T , Gaudet D , Hudson T J , Daly M , Groop L , Lander E S . The common PPARγ Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet. 2000;26:76–80. [PubMed: 10973253]
- 184.
- Chutkow W A , Samuel V , Hansen P A , Pu J , Valdivia C R , Makielski J C , Burant C F . Disruption of Sur2-containing K(ATP) channels enhances insulin-stimulated glucose uptake in skeletal muscle. Proc Natl Acad Sci U S A. 2001;98:11760–4. [PMC free article: PMC58803] [PubMed: 11562480]
- 185.
- Tammur J , Prades C , Arnould I , Rzhetsky A , Hutchinson A , Adachi M , Schuetz J D , Swoboda K J , Ptacek L J , Rosier M , Dean M , Allikmets R . Two new genes from the human ATP-binding cassette transporter superfamily, ABCC11 and ABCC12, tandemly duplicated on chromosome 16q12. Gene. 2001;273:89–96. [PubMed: 11483364]
- 186.
- Bera T K , Lee S , Salvatore G , Lee B , Pastan I . MRP8, a new member of ABC transporter superfamily, identified by EST database mining and gene prediction program, is highly expressed in breast cancer. Mol Med. 2001;7:509–16. [PMC free article: PMC1950066] [PubMed: 11591886]
- 187.
- Yabuuchi H , Shimizu H , Takayanagi S , Ishikawa T . Multiple splicing variants of two new human ATP-binding cassette transporters, ABCC11 and ABCC12. Biochem Biophys Res Commun. 2001;288:933–9. [PubMed: 11688999]
- 188.
- Turriziani O , Schuetz JD , Focher F , Scagnolari C , Sampath J , Adachi M , Bambacioni F , Riva E , Antonelli G . Impaired 2′,3′-dideoxy-3′-thiacytidine accumulation in T-lymphoblastoid cells as a mechanism of acquired resistance independent of multidrug resistant protein 4 with a possible role for ATP-binding cassette C11. Biochem J. 2002;368:325–332. [PMC free article: PMC1222956] [PubMed: 12133003]
- 189.
- Mosser J , Douar A M , Sarde C O , Kioschis P , Feil R , Moser H , Poustka A M , Mandel J L , Aubourg P . Putative X-linked adrenoleukodystrophy gene shares unexpected homology with ABC transporters. Nature. 1993;361:726–30. [PubMed: 8441467]
- 190.
- Kemp S , Pujol A , Waterham H R , van Geel B M , Boehm C D , Raymond G V , Cutting G R , Wanders R J , Moser H W . ABCD1 mutations and the X-linked adrenoleukodystrophy mutation database: role in diagnosis and clinical correlations. Hum Mutat. 2001;18:499–515. [PubMed: 11748843]
- 191.
- Holmberg B H , Hagg E , Hagenfeldt L . Adrenomyeloneuropathy—report on a family. J Intern Med. 1991;230:535–8. [PubMed: 1660916]
- 192.
- Kutsche K , Ressler B , Katzera H G , Orth U , Gillessen-Kaesbach G , Morlot S , Schwinger E , Gal A . Characterization of breakpoint sequences of five rearrangements in L1CAM and ABCD1 (ALD) genes. Hum Mutat. 2002;19:526–35. [PubMed: 11968085]
- 193.
- Corzo D , Gibson W , Johnson K , Mitchell G , LePage G , Cox G F , Casey R , Zeiss C , Tyson H , Cutting G R , Raymond G V , Smith K D , Watkins P A , Moser A B , Moser H W , Steinberg S J . Contiguous deletion of the X-linked adrenoleukodystrophy gene (ABCD1) and DXS1357E: a novel neonatal phenotype similar to peroxisomal biogenesis disorders. Am J Hum Genet. 2002;70:1520–31. [PMC free article: PMC419992] [PubMed: 11992258]
- 194.
- Aubourg P , Mosser J , Douar A M , Sarde C O , Lopez J , Mandel J L . Adrenoleukodystrophy gene: unexpected homology to a protein involved in peroxisome biogenesis. Biochimie. 1993;75:293–302. [PubMed: 8507690]
- 195.
- Poulos A , Gibson R , Sharp P , Beckman K , Grattan-Smith P . Very long chain fatty acids in X-linked adrenoleukodystrophy brain after treatment with Lorenzo's oil. Ann Neurol. 1994;36:741–6. [PubMed: 7979219]
- 196.
- Forss-Petter S , Werner H , Berger J , Lassmann H , Molzer B , Schwab M H , Bernheimer H , Zimmermann F , Nave K A . Targeted inactivation of the X-linked adrenoleukodystrophy gene in mice. J Neurosci Res. 1997;50:829–43. [PubMed: 9418970]
- 197.
- Yamada T , Shinnoh N , Kondo A , Uchiyama A , Shimozawa N , Kira J , Kobayashi T . Very-long-chain fatty acid metabolism in adrenoleukodystrophy protein- deficient mice. Cell Biochem Biophys. 2000;32:239–46. [PubMed: 11330052]
- 198.
- Pujol A , Hindelang C , Callizot N , Bartsch U , Schachner M , Mandel J L . Late onset neurological phenotype of the X-ALD gene inactivation in mice: a mouse model for adrenomyeloneuropathy. Hum Mol Genet. 2002;11:499–505. [PubMed: 11875044]
- 199.
- Shani N , Valle D . A Saccharomyces cerevisiae homolog of the human adrenoleukodystrophy transporter is a heterodimer of two half ATP-binding cassette transporters. Proc Natl Acad Sci U S A. 1996;93:11901–11906. [PMC free article: PMC38156] [PubMed: 8876235]
- 200.
- Verleur N , Hettema E H , van Roermund C W , Tabak H F , Wanders R J . Transport of activated fatty acids by the peroxisomal ATP-binding-cassette transporter Pxa2 in a semi-intact yeast cell system. Eur J Biochem. 1997;249:657–61. [PubMed: 9395310]
- 201.
- Zolman B K , Silva I D , Bartel B . The Arabidopsis pxa1 mutant is defective in an ATP-binding cassette transporter-like protein required for peroxisomal fatty acid β-oxidation. Plant Physiol. 2001;127:1266–78. [PMC free article: PMC129294] [PubMed: 11706205]
- 202.
- Lombard-Platet G , Savary S , Sarde C O , Mandel J L , Chimini G . A close relative of the adrenoleukodystrophy (ALD) gene codes for a peroxisomal protein with a specific expression pattern. Proc Natl Acad Sci U S A. 1996;93:1265–9. [PMC free article: PMC40068] [PubMed: 8577752]
- 203.
- Savary S , Troffer-Charlier N , Gyapay G , Mattei M G , Chimini G . Chromosomal localization of the adrenoleukodystrophy-related gene in man and mice. Eur J Hum Genet. 1997;5:99–101. [PubMed: 9195160]
- 204.
- Broccardo C , Troffer-Charlier N , Savary S , Mandel J L , Chimini G . Exon organisation of the mouse gene encoding the adrenoleukodystrophy related protein (ALDRP). Eur J Hum Genet. 1998;6:638–41. [PubMed: 9887385]
- 205.
- Netik A , Forss-Petter S , Holzinger A , Molzer B , Unterrainer G , Berger J . Adrenoleukodystrophy-related protein can compensate functionally for adrenoleukodystrophy protein deficiency (X-ALD): implications for therapy. Hum Mol Genet. 1999;8:907–13. [PubMed: 10196381]
- 206.
- Fourcade S , Savary S , Albet S , Gauthe D , Gondcaille C , Pineau T , Bellenger J , Bentejac M , Holzinger A , Berger J , Bugaut M . Fibrate induction of the adrenoleukodystrophy-related gene (ABCD2): promoter analysis and role of the peroxisome proliferator-activated receptor PPARα Eur J Biochem. 2001;268:3490–500. [PubMed: 11422379]
- 207.
- Gartner J , Moser H , Valle D . Mutations in the 70K peroxisomal membrane protein gene in Zellweger syndrome. Nat Genet. 1992;1:16–23. [PubMed: 1301993]
- 208.
- Paton B C , Heron S E , Nelson P V , Morris C P , Poulos A . Absence of mutations raises doubts about the role of the 70-kD peroxisomal membrane protein in Zellweger syndrome. Am J Hum Genet. 1997;60:1535–9. [PMC free article: PMC1716138] [PubMed: 9199576]
- 209.
- Shani N , Jimenez-Sanchez G , Steel G , Dean M , Valle D . Identification of a fourth half ABC transporter in the human peroxisomal membrane. Hum Mol Genet. 1997;6:1925–31. [PubMed: 9302272]
- 210.
- Holzinger A , Kammerer S , Roscher A A . Primary structure of human PMP69, a putative peroxisomal ABC-transporter. Biochem Biophys Res Commun. 1997;237:152–7. [PubMed: 9266848]
- 211.
- Holzinger A , Roscher A A , Landgraf P , Lichtner P , Kammerer S . Genomic organization and chromosomal localization of the human peroxisomal membrane protein-1-like protein (PXMP1-L) gene encoding a peroxisomal ABC transporter. FEBS Lett. 1998;426:238–42. [PubMed: 9599016]
- 212.
- Diriong S , Salehzada T , Bisbal C , Martinand C , Taviaux S . Localization of the ribonuclease L inhibitor gene (RNS4I), a new member of the interferon-regulated 2-5A pathway, to 4q31 by fluorescence in situ hybridization. Genomics. 1996;32:488–90. [PubMed: 8838820]
- 213.
- Bisbal C , Martinand C , Silhol M , Lebleu B , Salehzada T . Cloning and characterization of a RNAse L inhibitor. A new component of the interferon-regulated 2-5A pathway. J Biol Chem. 1995;270:13308–13317. [PubMed: 7539425]
- 214.
- Aubry F , Mattei M G , Barque J P , Galibert F . Chromosomal localization and expression pattern of the RNase L inhibitor gene. FEBS Lett. 1996;381:135–9. [PubMed: 8641422]
- 215.
- Zimmerman C , Klein K C , Kiser P K , Singh A R , Firestein B L , Riba S C , Lingappa J R . Identification of a host protein essential for assembly of immature HIV-1 capsids. Nature. 2002;415:88–92. [PubMed: 11780123]
- 216.
- Richard M , Drouin R , Beaulieu A D . ABC50, a novel human ATP-binding cassette protein found in tumor necrosis factor-α-stimulated synoviocytes. Genomics. 1998;53:137–45. [PubMed: 9790762]
- 217.
- Savary S , Denizot F , Luciani M , Mattei M , Chimini G . Molecular cloning of a mammalian ABC transporter homologous to Drosophila white gene. Mamm Genome. 1996;7:673–6. [PubMed: 8703120]
- 218.
- Croop J M , Tiller G E , Fletcher J A , Lux M L , Raab E , Goldenson D , Son D , Arciniegas S , Wu R L . Isolation and characterization of a mammalian homolog of the Drosophila white gene. Gene. 1997;185:77–85. [PubMed: 9034316]
- 219.
- Annilo T , Tammur J , Hutchinson A , Rzhetsky A , Dean M , Allikmets R . Human and mouse orthologs of a new ATP-binding cassette gene, ABCG4. Cytogenet Cell Genet. 2001;94:196–201. [PubMed: 11856881]
- 220.
- Schmitz G , Langmann T , Heimerl S . Role of ABCG1 and other ABCG family members in lipid metabolism. J Lipid Res. 2001;42:1513–1520. [PubMed: 11590207]
- 221.
- Langmann T , Porsch-Ozcurumez M , Unkelbach U , Klucken J , Schmitz G . Genomic organization and characterization of the promoter of the human ATP-binding cassette transporter-G1 (ABCG1) gene. Biochim Biophys Acta. 2000;1494:175–80. [PubMed: 11072082]
- 222.
- Allikmets R , Schriml L M , Hutchinson A , Romano-Spica V , Dean M . A human placenta-specific ATP-binding cassette gene (ABCP) on chromosome 4q22 that is involved in multidrug resistance. Cancer Res. 1998;58:5337–5339. [PubMed: 9850061]
- 223.
- Miyake K , Mickley L , Litman T , Zhan Z , Robey R , Cristensen B , Brangi M , Greenberger L , Dean M , Fojo T , Bates S E . Molecular cloning of cDNAs which are highly overexpressed in mitoxantrone-resistant cells: demonstration of homology to ABC transport genes. Cancer Res. 1999;59:8–13. [PubMed: 9892175]
- 224.
- Doyle L A , Yang W , Abruzzo L V , Krogmann T , Gao Y , Rishi A K , Ross D D . A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc Natl Acad Sci U S A. 1998;95:15665–70. [PMC free article: PMC28101] [PubMed: 9861027]
- 225.
- Ross D D , Yang W , Abruzzo L V , Dalton W S , Schneider E , Lage H , Dietel M , Greenberger L , Cole S P , Doyle L A . Atypical multidrug resistance: breast cancer resistance protein messenger RNA expression in mitoxantrone-selected cell lines. J Natl Cancer Inst. 1999;91:429–33. [PubMed: 10070941]
- 226.
- Honjo Y , Hrycyna C A , Yan Q W , Medina-Perez W Y , Robey R W , van de Laar A , Litman T , Dean M , Bates S E . Acquired mutations in the MXR/BCRP/ABCP gene alter substrate specificity in MXR/BCRP/ABCP-overexpressing cells. Cancer Res. 2001;61:6635–9. [PubMed: 11559526]
- 227.
- Litman T , Brangi M , Hudson E , Fetsch P , Abati A , Ross D D , Miyake K , Resau J H , Bates S E . The multidrug-resistant phenotype associated with overexpression of the new ABC half-transporter, MXR (ABCG2). J Cell Sci. 2000;113:2011–21. [PubMed: 10806112]
- 228.
- Brangi M , Litman T , Ciotti M , Nishiyama K , Kohlhagen G , Takimoto C , Robey R , Pommier Y , Fojo T , Bates S E . Camptothecin resistance: role of the ATP-binding cassette (ABC), mitoxantrone-resistance half-transporter (MXR), and potential for glucuronidation in MXR-expressing cells. Cancer Res. 1999;59:5938–46. [PubMed: 10606239]
- 229.
- Zhou S , Schuetz J D , Bunting K D , Colapietro A M , Sampath J , Morris J J , Lagutina I , Grosveld G C , Osawa M , Nakauchi H , Sorrentino B P . The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med. 2001;7:1028–34. [PubMed: 11533706]
- 230.
- Rocchi E , Khodjakov A , Volk E L , Yang C H , Litman T , Bates S E , Schneider E . The product of the ABC half-transporter gene ABCG2 (BCRP/MXR/ABCP) is expressed in the plasma membrane. Biochem Biophys Res Commun. 2000;271:42–6. [PubMed: 10777678]
- 231.
- Jonker J W , Smit J W , Brinkhuis R F , Maliepaard M , Beijnen J H , Schellens J H , Schinkel A H . Role of breast cancer resistance protein in the bioavailability and fetal penetration of topotecan. J Natl Cancer Inst. 2000;92:1651–6. [PubMed: 11036110]
- 232.
- de Bruin M , Miyake K , Litman T , Robey R , Bates S E . Reversal of resistance by GF120918 in cell lines expressing the ABC half-transporter, MXR. Cancer Lett. 1999;146:117–26. [PubMed: 10656616]
- 233.
- Rabindran S K , He H , Singh M , Brown E , Collins K I , Annable T , Greenberger L M . Reversal of a novel multidrug resistance mechanism in human colon carcinoma cells by fumitremorgin C. Cancer Res. 1998;58:5850–5858. [PubMed: 9865745]
- 234.
- Rabindran S K , Ross D D , Doyle L A , Yang W , Greenberger L M . Fumitremorgin C reverses multidrug resistance in cells transfected with the breast cancer resistance protein. Cancer Res. 2000;60:47–50. [PubMed: 10646850]
- 235.
- Engel T , Lorkowski S , Lueken A , Rust S , Schluter B , Berger G , Cullen P , Assmann G . The human ABCG4 gene is regulated by oxysterols and retinoids in monocyte-derived macrophages. Biochem Biophys Res Commun. 2001;288:483–8. [PubMed: 11606068]
- 236.
- Yoshikawa M , Yabuuchi H , Kuroiwa A , Kegami Y , Sai Y , Tamai I , Tsuji A , Matsuda Y , Yoshida H , Ishikawa T . Molecular and cytogenetic characterization of the mouse ATP-binding cassette transporter Abcg4. Gene. 2002;293:67–75. [PubMed: 12137944]
- 237.
- Shulenin S , Schriml L M , Remaley A T , Fojo S , Brewer B , Allikmets R , Dean M . An ATP-binding cassette gene (ABCG5) from the ABCG (White) gene subfamily maps to human chromosome 2p21 in the region of the sitosterolemia locus. Cytogenet Cell Genet. 2001;92:204–8. [PubMed: 11435688]
- 238.
- Berge K E , Tian H , Graf G A , Yu L , Grishin N V , Schultz J , Kwiterovich P , Shan B , Barnes R , Hobbs H H . Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science. 2000;290:1771–5. [PubMed: 11099417]
- 239.
- Lee M H , Lu K , Hazard S , Yu H , Shulenin S , Hidaka H , Kojima H , Allikmets R , Sakuma N , Pegoraro R , ivastava A K , Salen G , Dean M , Patel S B . Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat Genet. 2001;27:79–83. [PMC free article: PMC1350991] [PubMed: 11138003]
- 240.
- Bhattacharyya A K , Connor W E . β-Sitosterolemia and xanthomatosis. A newly described lipid storage disease in two sisters. J Clin Invest. 1974;53:1033–43. [PMC free article: PMC333088] [PubMed: 4360855]
- 241.
- Salen G , Shefer S , Nguyen L , Ness G C , Tint G S , Batta A K . Sitosterolemia. Subcell Biochem. 1997;28:453–76. [PubMed: 9090303]
- 242.
- Gregg R E , Connor W E , Lin D S , Brewer H B Jr. Abnormal metabolism of shellfish sterols in a patient with sitosterolemia and xanthomatosis. J Clin Invest. 1986;77:1864–72. [PMC free article: PMC370545] [PubMed: 3711338]
- 243.
- Patel S B , Salen G , Hidaka H , Kwiterovich P O , Stalenhoef A F , Miettinen T A , Grundy S M , Lee M H , Rubenstein J S , Polymeropoulos M H , Brownstein M J . Mapping a gene involved in regulating dietary cholesterol absorption. The sitosterolemia locus is found at chromosome 2p21. J Clin Invest. 1998;102:1041–4. [PMC free article: PMC508970] [PubMed: 9727073]
- 244.
- Remaley A T , Bark S , Walts A D , Freeman L , Shulenin S , Annilo T , Elgin E , Rhodes H E , Joyce C , Dean M , Santamarina-Fojo S , Brewer H B . Comparative genome analysis of potential regulatory elements in the ABCG5–ABCG8 gene cluster. Biochem Biophys Res Commun. 2002;295:276–82. [PubMed: 12150943]
- 245.
- Lu K , Lee M H , Hazard S , Brooks-Wilson A , Hidaka H , Kojima H , Ose L , Stalenhoef A F , Mietinnen T , Bjorkhem I , Bruckert E , Pandya A , Brewer H B Jr., Salen G , Dean M , Srivastava A , Patel S B . Two genes that map to the STSL locus cause sitosterolemia: genomic structure and spectrum of mutations involving sterolin-1 and sterolin- 2, encoded by ABCG5 and ABCG8, respectively. Am J Hum Genet. 2001;69:278–90. [PMC free article: PMC1201544] [PubMed: 11452359]
- 246.
- Lam C W , Cheng A W , Tong S F , Chan Y W . Novel donor splice site mutation of ABCG5 gene in sitosterolemia. Mol Genet Metab. 2002;75:178–80. [PubMed: 11855938]
- 247.
- Heimer S , Langmann T , Moehle C , Mauerer R , Dean M , Beil F U , Von Bergmann K , Schmitz G . Mutations in the human ATP-binding cassette transporters ABCG5 and ABCG8 in sitosterolemia. Hum Mutat. 2002;20:151. [PubMed: 12124998]
- 248.
- Berge K E , von Bergmann K , Lutjohann D , Guerra R , Grundy S M , Hobbs H H , Cohen J C . Heritability of plasma noncholesterol sterols and relationship to DNA sequence polymorphism in ABCG5 and ABCG8. J Lipid Res. 2002;43:486–94. [PubMed: 11893785]
- 249.
- Hubacek J A , Berge K E , Cohen J C , Hobbs H H . Mutations in ATP-cassette binding proteins G5 (ABCG5) and G8 (ABCG8) causing sitosterolemia. Hum Mutat. 2001;18:359–60. [PubMed: 11668628]
- 250.
- Bateman A , Birney E , Durbin R , Eddy S R , Finn R D , Sonnhammer E L . Pfam 3.1: 1313 multiple alignments and profile HMMs match the majority of proteins. Nucleic Acids Res. 1999;27:260–2. [PMC free article: PMC148151] [PubMed: 9847196]
- 251.
- Eddy S R . Profile hidden Markov models. Bioinformatics. 1998;14:755–63. [PubMed: 9918945]
- 252.
- Eddy S R . Multiple alignment using hidden Markov models. ISMB. 1995;3:114–20. [PubMed: 7584426]
- 253.
- Saitou N , Nei M . The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–25. [PubMed: 3447015]
- 254.
- Zuckerkandl E, Pauling L. Evolutionary divergence and convergence in proteins. In: BA Vogel (editor). Evolving genes and proteins. New York: Academic Press, pp. 97-166; 1965.
- 255.
- Jones D T , Taylor W R , Thornton J M . The rapid generation of mutation data matrices from protein sequences. Comput Appl Biosci. 1992;8:275–82. [PubMed: 1633570]
- 256.
- Zwarts K Y , Clee S M , Zwinderman A H , Engert J C , Singaraja O , Loubser J M , James E , Roomp K , Hudson T J , Jukema J W , Kastelein J J , Hayden M R . ABCA1 regulatory variants influence coronary artery disease independent of effects on plasma lipid levels. Clin Genet. 2002;61:115–125. [PubMed: 11940086]
- 257.
- Szakacs G , Langmann T , Ozvegy C , Orso E , Schmitz G , Varadi A , Sarkadi B . Characterization of the ATPase cycle of human ABCA1: implications for its function as a regulator rather than an active transporter. Biochem Biophys Res Commun. 2001;288:1258–1264. [PubMed: 11700048]
- 258.
- Vulevic B , Chen Z , Boyd J T , Davis W Jr, Walsh E S , Belinsky M G , Tew K D . Cloning and characterization of human adenosine 5'-triphosphate-binding cassette, sub-family A, transporter 2 (ABCA2). Cancer Res. 2001;61:3339–3347. [PubMed: 11309290]
- 259.
- Decottignies A , Goffeau A . Complete inventory of the yeast ABC proteins. Nat Genet. 1997;15:137–145. [PubMed: 9020838]
- 260.
- Michaelis S , Berkower C . Sequence comparison of yeast ATP binding cassette (ABC) proteins. Cold Spring Harbor Symposium. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1995. [PMC free article: PMC358499] [PubMed: 8824403]
Created: November 18, 2002.
- Introduction to ABC Protein and Gene Organization
- Nomenclature
- Overview of Human ABC Gene Subfamilies
- ABC Genes and Human Genetic Disease
- Mouse Knockouts
- Multidrug Resistance and Cancer Therapy
- Phylogenetic Analysis of Human ABC Genes
- Mouse ABC Genes
- Drosophila ABC Genes
- ABCA Genes
- ABCB Genes
- ABCC Genes
- ABCD Genes
- ABCE Genes
- ABCF Genes
- ABCG Genes
- References
- NLM CatalogRelated NLM Catalog Entries
- OMIMRelated OMIM records
- PMCPubMed Central citations
- PubMedLinks to PubMed
- GeneLocus Links
- The Human ATP-Binding Cassette (ABC) Transporter SuperfamilyThe Human ATP-Binding Cassette (ABC) Transporter Superfamily
- putative aminopeptidase, partial [Trypanosoma cruzi]putative aminopeptidase, partial [Trypanosoma cruzi]gi|2583639954|gb|WNO13686.1|Protein
- Trypanosoma cruzi trypanothione reductase (TR) gene, partial cds.Trypanosoma cruzi trypanothione reductase (TR) gene, partial cds.PopSet: 2583640097PopSet
- LOC125078067 [Homo sapiens]LOC125078067 [Homo sapiens]Gene ID:125078067Gene
- J5X95_RS20895 [Bacillus amyloliquefaciens]J5X95_RS20895 [Bacillus amyloliquefaciens]Gene ID:75096023Gene
Your browsing activity is empty.
Activity recording is turned off.
See more...